摘要
茶树(Camellia sinensis(L.) O. Ktze.) 是一种世界广泛种植且具有经济效益的叶用作物。因为具有特定叶色的茶叶的特征化学成分与茶叶质量密切相关,因而茶树叶色多样性的研究一直以来被茶业界广为关注。本研究以黄山群体中花白茶树单株为研究材料,发现花白茶树不同于温敏型白化茶树和光敏型白化茶树随着温度和光照强度变化而变化,经过修剪发现花白性状依然存在。于是展开花白茶树中内含成分、组织结构、基因表达差异和蛋白表达差异等研究,取得以下研究结果:
(1)通过内含物质的含量比较分析发现,相对于正常叶片,花白叶片的叶绿素含量显著降低,咖啡碱含量略有增长;相较于正常叶片,花白叶片中总儿茶素含量减少了52.66%,而总游离氨基酸含量增加了52.15%,其中花白叶片的茶氨酸含量增长了62.73%。通过石蜡切片发现花白叶片栅栏组织细胞排列疏松,组织结构的疏松程度显著低于正常叶片,并且观察花白叶片的叶绿体超微结构发现花白叶片的叶绿体发育异常,没有明显的类囊体且存在空腔。
(2)通过对花白叶片与正常叶片转录组学分析,共获得3431个差异表达基因。针对茶树特征性代谢中茶氨酸和多酚生物合成相关基因进行分析,发现花白叶片在儿茶素代谢途径中的相关基因低于正常叶片,而在茶氨酸代谢途径中的基因趋势相反。通过分析还发现,花白叶片中叶绿素生物合成代谢途径、光合作用、叶绿体发育和生物合成中差异基因的表达模式受到影响。花白叶片中光合作用的光捕获天线蛋白基因(LHC)表达均显著低于正常叶片,合成叶绿素途径相关的差异基因表达也低于正常。
(3)通过对花白茶树中花白叶片和正常叶片的蛋白组分析,一共获得95个差异蛋白。经过KEGG富集分析,结果发现花白茶树在氧化磷酸化,类黄酮生物途径和苯丙素生物合成等重要通路发生了显著变化,同时在光合作用、嘌呤代谢等代谢途径受到影响。
(4)我们在对差异基因进行筛选时发现了Clp蛋白酶复合体与叶绿体发育、植物叶色有关。以花白叶片和正常叶片为模板,分别扩增CsClpP5基因。经序列分析发现,从花白叶片中克隆的CsClpP5基因,全长序列为909bp,编码303个氨基酸。然而正常叶片中扩增的CsClpP5基因全长903bp,编码301个氨基酸。经过序列比对发现了两个氨基酸位点的突变,而经过磷酸位点的分析发现该突变位点的改变了ClpP5的磷酸位点。由此我们推测CsClpP5突变,可能与茶树产生花白表型有关。
关键词:茶树;白化;转录组;蛋白组;CsClpP5基因
1 文献综述
1.1白化茶树的研究进展
作为茶树的发源地,中国的茶树资源丰富,其中白化茶树因独特茶味,包括高鲜味和低涩味,使得白化茶树从茶树(Camellia sinesnis)中脱颖而出。根据不同的环境因素,可以将白化茶树分为光敏感型白化茶树、温度敏感型白化茶树和生态不敏感型白化茶树[1]。
1.1.1光敏型白化茶树
黄金芽、白鸡冠、御金香为光敏型白化茶树的主要品种,叶片在强光照(> 15000 lx)条件下黄化,但随着光照强度的降低或遮荫条件下逐渐恢复绿色[2-4]。温敏型茶树的白化一般出现在晚春寒潮期,这种特殊的气候现象导致气温明显低于年平均气温,有时甚至会降至10℃以下[5]。黄金芽的白化叶片大大减少了叶绿素的含量,经检测黄金芽白化叶片的叶绿素a含量减少了96%以及叶绿素b降低了77%。黄金芽的超微结构显示白化叶片的叶绿体在前质体期被抑制,没有基粒结构,取而代之的是普遍存在的嗜锇颗粒。黄金芽的白化叶片比非白化叶片含有茶氨酸、谷氨酸、谷氨酰胺、天冬氨酸分别升高31.0、39.8、50.8、62.2%[2]。叶绿体类囊体膜在白鸡冠的白化叶片中受到损坏。研究发现白鸡冠的白化叶片(不遮荫)与对照(遮荫)相比,儿茶素含量显著降低,主要集中在表没食子儿茶素、儿茶素和表儿茶素等非酯型儿茶素[3]。御金香白化叶片的叶肉细胞中叶绿体少于遮荫后的叶片,且叶绿体的形态异常。白化叶片中EGCG、EGC、ECG与EC均低于遮荫后的叶片,其中槲皮素与山柰醇在御金香白化叶片中有所积累,而茶氨酸在遮荫后的更高含量[4]。
1.1.2 温敏型白化茶树
安吉白茶、千年雪、小雪芽以及黄山白茶1号是温敏型白化茶树的主要扦插对象,它们在低温(< 20℃)下萌发出雪白嫩叶,在高温(22至25℃)下恢复标准叶色[6, 7]。安吉白茶在白化期间叶绿体的发育不完全,类囊体结构中的基粒模糊。白化严重时期的叶肉细胞中并未发现叶绿体。安吉白茶在白化期的茶氨酸和胡萝卜素高于绿叶期[8]。千年雪的抗逆性比安吉白茶的强[9],氨基酸和多酚含量与安吉白茶所含的差异不大[10]。与小雪芽的绿叶组织比较,发现小雪芽的白化叶片所含的叶绿素和类胡萝卜素显著不足,叶绿体严重破坏。此外,小雪芽的白化叶片中与品质有关的次生代谢物的积累也发生了改变,总氨基酸、茶氨酸和谷氨酸含量显著增加,生物碱、儿茶素和多酚含量显著降低[11]。黄山白茶1号在白化时期所含的叶绿素含量低而游离氨基酸含量增加。其中黄山白茶1号白化阶段总氨基酸的含量是复绿阶段的2.6倍[12]。
1.1.3 生态不敏感型白化茶树
不同于光敏型与温敏型白化茶树受环境所影响,在不考虑环境的情况下,产生稳定的、且可遗传的杂色叶片称为生态不敏感型白化茶树。其中主要包括炎陵银边茶、黄金斑、银红九号等变异体[13-15]。变异的炎陵银边茶存在复色的表型,研究中发现白化部分中叶绿体的基粒发育不良,并有许多的空泡结构存在。其内含成分咖啡碱、表没食子儿茶素没食子酸酯和没食子儿茶素没食子酸酯以及表儿茶素没食子酸酯比正常组织含量低[13]。黄金斑的研究较少,是黄金芽的白化变种,黄金斑茶树在视觉上构成了黄绿相间的复色状态[14]。黄化的英红九号中叶绿体缺乏结构良好的类囊体膜,类囊体肿胀,且叶绿体的形态异常[16]。
1.1.4 白化茶树的研究前景
与其他植物最大的不同在于,白化茶树具有人类所追求的品质特征。茶氨酸和儿茶素是茶树中最重要的次生代谢物,它们不仅在调节碳氮代谢方面有显著的作用,并且对茶的风味也有贡献,并与人体健康和药用价值呈正相关[17, 18]。白化茶树的白化期L-茶氨酸含量是常规茶树品种的0.5到2倍,但总儿茶素含量只有常规茶树品种的一半,导致多酚和氨基酸的比例被显著降低,这使得白化茶树加工的绿茶不含苦味,而具有非凡的鲜味[19]。目前,由于白化突变,白化茶树的种植收入是普通品种的十倍[20]。与此同时,白化茶树中白化色泽不一,存在白色、黄色、复色等独特的色系。与天竺葵、洒金珊瑚等植株类似,都具有观赏性价值。白化茶树可以作为茶园观赏品种,可应用于园林设计、规划等[21]。因此,白化茶树在饮用、食用、园林设计等方面中具有极高的开发潜力,广泛的推广培育并且充分的利用把握白化品种茶树资源有着深远的意义。
1.2白化植物的分子研究
白化(也称为白化病)在植物界中是一种常见的变异,其中大量的白化突变体中在拟南芥[22]、玉米[23]、棉花突变体[24]和水稻突变体[25]以及番茄突变体[26]等等发现。叶片颜色变化的发生主要受遗传和环境的影响。有研究表明植物叶色的变化是由一个或多个因素引起的,包括基因型、环境、激素失衡、核质体基因组不协调性、质体DNA缺失、与叶绿素生物合成相关的基因突变以及叶绿素生物合成途径的代谢阻断[27]。质体基因[28-30]或大约300个核基因[31]的突变都可以产生白化表型。而这些基因的突变会直接或者间接地调节叶绿体的结构、叶绿体的生物合成和相关影响叶色的代谢过程[32, 33]。对于大多数作物来说,白化症状由于破坏了光合器官(例如叶绿体),导致碳和氮代谢失衡,造成产量低、不育等不良影响[34, 35]。此外,环境因素的变化,包括温度[36]、光照[37]和矿物质元素[38],也会导致叶片颜色的变化。由于叶色的变化是一个或多个因素造成的,因此这在研究叶色变化的内在机制带来了困难。
1.2.1与叶绿体结构发育相关的基因突变
对于植物来说叶绿体是一个重要的细胞器官,也作为植物的一个重要特征。叶绿体是光合作用的场所,同时也是脂肪酸生物合成、硝酸盐同化和氨基酸生物合成代谢途径的场所,并且在植物发育过程中起着重要的作用。
Clp蛋白酶(酪蛋白水解酶)是依赖ATP型的蛋白水解酶,主要定位于质体,尤其是在叶绿体的基质中。在拟南芥中,Clp蛋白酶复合体是由多个不同家族的Clp蛋白酶所组成,包括5个ClpP亚基(ClpP1、ClpP3、ClpP4、ClpP5和ClpP6)和4个ClpR亚基(ClpR1到ClpR4)组成一个325-350 kDa的四聚体蛋白酶核心,2个ClpC (CIpC1和ClpC2)和1个ClpD伴侣蛋白(adaptor ClpS1),以及2个植物特异性辅助蛋白,ClpT1和ClpT2。Clp蛋白酶复合体主要的功能是清除无法挽回的、损伤的蛋白质进而能调节植物的蛋白质代谢。同时,为了保证细胞的正常生理机能,Clp蛋白酶复合体还可以在代谢过程中通过调节限速酶的水平来控制新陈代谢过程。[39]。基质Clp蛋白酶通过降解关键调节的蛋白或酶在叶绿体发育分化中起到不可或缺的作用。
Clp蛋白酶复合体是质体基质中蛋白降解的主要机制[40]。目前有些研究揭示了Clp蛋白酶复合体的降解底物,而这些底物中有相当一部分是正常叶绿体代谢所必需的,包括参与产生必需的四吡咯和类异戊二烯如叶绿素和类胡萝卜素代谢途径中参与的酶。核编码的Clp蛋白酶基因表达量降低或者完全缺失可导致拟南芥的叶色发生变化。通过反义RNA技术降低ClpP4和ClpP6表达量,能使得拟南芥变为淡绿色植株[41, 42]。借助甲基磺酸乙酯(EMS)诱变或者插入T-DNA降低ClpR1蛋白含量(几乎检测不到的水平)可导致淡绿色突变体[43, 44]。有研究表明叶绿体Clp蛋白酶复合体中的ClpP5亚基蛋白酶基因突变导致基因功能缺失会导致玉米幼苗叶片黄化[45]。
拟南芥突变体skl1-8是单个隐形核基因(莽草酸激酶)突变表现为白化的表型,其特征是色素缺陷型。检测skl1-8的叶绿素含量结果发现显著降低并且发现叶绿体发育受阻结构受损。在拟南芥突变体skl1-8中光合蛋白水平大幅降低,以及光合作用、色素代谢和叶绿体相关的基因表达受到抑制。同样地,在skl1-8突变体中参与生长素转运和反应的相关基因被抑制,表明SKL1在叶绿体发育过程中参与生长素相关的通路[46]。在拟南芥中鉴定出一个突变体发现突变的基因编码一个质体靶向的延伸因子G(SCO1)。该因子在胚胎发生过程中对质体的发育是必不可少的,因为编码序列中的两个T-DNA插入突变(sco1-2和sco1-3)导致了胚胎致死的表型。此外,SCO1的一个点突变等位基因(sco1-1)和一个T-DNA插入的等位基因(sco1-4)表现出条件性幼苗致死表型。这些等位基因的幼苗由于叶绿体发育不良而表现出子叶和下胚轴白化,通常在萌发后不久死亡。但是当突变植株在添加蔗糖的培养基中发芽的时候,便可以从顶端分生组织产生具有光合作用的绿叶[47]。
1.2.2叶绿素生物代谢中的基因突变
植物白化症状的特征是在正常的绿色组织中缺乏叶绿素。近年来研究发现叶绿素相关的生物合成途径中至少有15步反应,而其中参与每一步的基因几乎已被检验确定。叶绿素合成途径大概为:谷氨酸(Glu) →5-氨基乙酰丙酸(ALA) → 原卟啉IX(Proto IX)→镁原卟啉IX(Mg- Proto IX)→ 叶绿素a(Chl a)→ 叶绿素b(Chl b)。参加叶绿素生物合成通路中相关的16个酶中任何一个基因产生突变都有可能导致植物的叶色变化。
镁原卟啉IX是叶绿素生物合成的中间产物,同时可通过信号通路抑制核转录的一种质体信号。在拟南芥中,这种信号通路需要GUN4基因。GUN4可以通过结合镁-螯合酶的产物和底物来进行增强镁-螯合酶的活性,并且GUN4可以经过调控镁原卟啉IX的合成或转运参与胞核信号转导。拟南芥中GUN4-1突变体产生为淡绿的表型,同时还抑制了叶绿素的积累。拟南芥GUN4-2突变体生长矮小,在标准的生长条件下表现为白化、黄化的表型[48]。在叶绿素生物合成途径中,其中需要在光条件下的一步反应是原叶绿素通过NADPH-原叶绿素氧化还原酶(POR)催化从而转化为叶绿素[49]。原卟啉IX是在血红素和叶绿素的生物合成途径之间最后一个也是普遍常见的中间产物,而镁的加入将这种分子导向叶绿素生物合成。一个镁原卟啉IX甲基转移酶(CHLM)缺失的拟南芥突变体表型失绿,会引起下游镁原卟啉IX的堵塞和叶绿素生物合成中间体的积累,并且发现CHLM基因对叶绿素的形成以及随后光系统I和II和细胞色素b6f复合物的形成至关重要[50]。在拟南芥[51]和水稻[52]中都有鉴定出POR基因,拟南芥中鉴定出3个POR基因其中包括PORA,PORB,和PORC,而在水稻中则存在2个POR基因(PORA,PORB)。拟南芥porc突变体的叶绿素浓度在强光下显著降低,并且porc的过度表达表现出对光氧化损伤越来越强的耐受性[53]。有研究发现在水稻叶色突变体FGL(褪色绿叶)中的PORB基因的第二外显子缺失了一个G碱基,从而产生了叶片的黄绿色表型[54]。
叶绿素的降解途径是随着叶绿素酶和金属螯合物质的相继除去植醇和镁而形成脱镁叶绿酸a (Pheide a),而随后最终将会被转化为无色无荧光的叶绿素分解代谢物。而一旦参与叶绿素降解途径的相关基因产生变化,植株便会产生叶色变化。在具有延迟叶片衰老的遗传变异突变体中,在叶片衰老过程中光合作用的结构的破坏会被部分或完全阻止。拟南芥不黄变的突变体nye1-1由于脱镁叶绿酸a加氧酶活性低,保留着50%的叶绿素。将突变体nye1-1中突变基因AtNYE1过表达进而增强脱镁叶绿酸a加氧酶活性,即可将拟南芥的表型呈现为淡黄色甚至白化[55]。有研究表明Sgr(stay green)基因编码叶绿体蛋白衰老的相关基因。而将Sgr基因过表达,发现转基因水稻会产生黄褐色的叶片[56]。
1.2.3与光合系统相关的基因突变
光系统II是存在于叶绿体类囊体膜上的一种色素复合蛋白,驱动电子从水转移到质体醌,并伴随着氧的进化。LPA2蛋白编码的是一种固有的类囊体膜蛋白,但却不是光系统II中一个完好的亚基。当LPA2基因发生突变,拟南芥lpa2-1突变体表现为浅绿色表型,并且生长速度明显降低。在lpa2-1突变体中研究发现叶绿素a结合蛋白CP43的合成量明显减少,在光系统II的复合物积累方面存在缺陷[57]。FZL是一个新的植物特异性成员。FZL蛋白是以叶绿体为靶点,与类囊体和包膜结合形成点状结构。将fzl基因敲除的突变体在叶绿体和类囊体形态上有异常,呈现散乱的颗粒状类囊体,并且叶色变淡以及花期延长。FZL-GFP过表达也导致类囊体组织缺陷[58]。类囊体膜的生物合成途径是植物光合作用不能或缺的通路。其中类囊体膜脂单半乳糖二酰基甘油(MGDG)构成了叶绿体膜脂的主要部分。而MGDG合成的最后一步是需要在MGDG合成酶在质体膜上完成。拟南芥MGD1突变体中MGDG合成酶功能受到严重的缺陷,突变的种子只有在蔗糖的存在下才会以白化症状的形式发芽。幼苗缺乏半乳脂质进而破坏了光合膜,导致光合能力和光自养生长完全受损[59]。
1.3 高通量测序和蛋白组学在白化茶树上的应用
组学技术是探索精确生命科学的关键方法,它整合和链接了多个组成领域的技术[60]。随着高通量技术的发展,大量组学研究实例在模型物种中的运行为白化茶树的研究提供了思绪。宛晓春团队[61]利用Illumina和PacBio等测序技术展示了茶树基因组的高质量序列。至少有64%的3.1 Gb的基因组组装是由重复序列组成的,其余的产生了33932个编码蛋白的高可信预测。根据计算,两个主要的世系中国种茶树CSS(Camellia sinensisvar. sinensis)和 阿萨姆种茶树CSA(Camellia sinensisvar. assamica)发现在38万到154万年前就发生了出现了分歧。相关证据表明,这些基因组复制事件以及随后的杂合复制对次级代谢物(儿茶素、茶氨酸和咖啡基因)的拷贝数有重大影响。与模式植物相比,大量基因组(3.1Gb基因组组装)、蛋白质和代谢物(大约700个生物活性化合物,由儿茶素、L-茶氨酸、咖啡因和挥发物组成)的表达和调控将更好地反映组学技术的完整性和相关性[61, 62]。
近年来,研究人员尝试在白化茶树中应用转录组学[8]、蛋白质组学[63, 64],极大地加快了研究进展。
光合作用所使用的光能来源于光合色素的吸收。叶绿素(主要的光合色素)的比例和含量对植物的表型和生理状态有重要影响。光照是在叶绿素生物合成途径中所必须的环境因子。叶绿素生物合成途径中,其中需要在光条件下的一步反应是原叶绿素通过NADPH-原叶绿素氧化还原酶(POR)催化从而转化为叶绿素[49]。在白化茶树中,白化或黄化叶片中叶绿素含量的显著下降总是伴随着POR及其编码基因表达的下降[65, 66]。通过计算PORA基因与叶绿素含量相关系数结果也表明两者呈现显著的正相关[8]。因此,白化茶树中POR基因的下调必须是启动白化症状的重要因素。此外,一些学者提出PORA蛋白的存在是为了建立前板层黄体的晶体结构[67]。PORA不仅作为一种催化酶在叶绿素生物合成途径中发挥作用,而且还作为一种自杀酶的形式来保护催化的产物叶绿素酸酯。另外,还有研究表明PORA蛋白在光下的分解是为了迅速形成类囊体膜[68]。因此,PORA的表达下调可能是导致叶绿体结构异常的原因之一,使得叶绿体结构发育不良,包括类囊体发育不良和基粒排列紊乱的结构异常现象。
植物将二氧化碳同化为碳水化合物的过程分为三个步骤,其中第一成为原初反应;二则是电子传递和光合磷酸化;第三是碳同化。第一步和第二步被称为“光反应”,必须要在光线下进行。相比之下,第三步(“暗反应”)通常不需要光线。光系统I (PSI)、光系统II (PSII)、细胞色素b6/f复合物和ATP酶(光反应涉及四个主要的蛋白质复合物)共同构成了自然界中一个微妙而巧妙的生物系统[69]。在温敏型白化茶树的安吉白茶中,所有差异积累蛋白都在与白化时期相对应的复绿期上调,表明随着叶绿体结构的恢复,光反应增强[63]。作为一种C3植物,茶树利用同化力通过卡尔文循环进行碳同化,许多酶参与了这一过程。例如,Rubisco(核酮糖-二磷酸羧化酶/加氧酶)等酶在植物光合作用中碳固定(暗反应)的反应中起着一个限速酶的作用。它们对于提高光合作用效率至关重要[70]。结果表明,与绿叶相比,黄化叶片中的Rubisco丰度显著降低[65],这表明除同化力不足外,碳代谢的抑制可能与气孔导度的关闭有关。在研究光敏型黄金芽的结果发现参与糖酵解途径的相关酶在失绿叶片中的表达水平较高,其中包括6-磷酸果糖激酶、丙酮酸激酶、磷酸甘油酸变位酶和果糖1,6-二磷酸酶[65],这表明不仅合成途径中的酶会影响白化茶树的碳同化,下游光合产物合成酶的表达也可以起到负反馈调节作用。
茶树中大多数的含氮化合物主要包括蛋白质、游离氨基酸和咖啡碱等化合物质。温敏型白化茶树安吉白茶在白化阶段中的茶氨酸合成相关基因表达受到抑制。与返绿后的正常叶片相比,处于白化阶段的白化叶片茶氨酸合成相关基因CsTS(茶氨酸合成酶)、CsGS(谷氨酰胺合成酶)和CsGOGAT(谷氨酸合成酶)的表达都受到了显著的抑制[71]。在研究光敏型白化茶树黄金芽的结果发现到编码铁氧还蛋白亚硝酸还原酶(CsFdNiR)以及硝酸还原酶(CsNR)、谷氨酰胺合成酶和茶氨酸合成酶的基因在白化叶片中的表达水平较低,而编码谷氨酸合成酶、谷氨酸脱氢酶(CsGDH)和天冬酰胺合成酶(CsAS)的基因表达水平较高[2]。而在温敏型白化茶树的黄山白茶1号中的研究表明:黄山白茶1号处于白化期间CsGS、CsSAMDC(S-腺苷甲硫氨酸脱羧酶)和CsADC(精氨酸脱羧酶)表达含量显著高于复绿阶段。乙胺是合成茶氨酸的底物之一,同样也是能够加强植物抗逆性的次生代谢产物。其中CsADC和CsSAMDC催化乙胺产生的关键酶,CsPAO(伯胺氧化酶)和CsCuAO(铜甲胺氧化酶)参与了乙胺的分解和调节。在黄山白茶1号的CsADC和CsSAMDC的表达水平显著高于黄山种。茶氨酸代谢途径的关键基因CsCuAO表达值在黄山白茶1号高于黄山种[12]。类似的结果也在光敏型白化茶树黄金芽发现,CsAO(乙胺水解酶)和CsSAMDC的表达水平显著高于迎霜[72]。
白化茶树中白化叶片的叶绿体和膜结构的异常发育抑制了光合作用,从而影响了糖代谢和碳同化。参与光合作用和碳水化合物代谢的二磷酸核酮糖羧化酶、己糖激酶、磷酸甘油酸变位酶、果糖激酶以及糖原磷酸化酶等相关基因在黄金芽白化的叶片中上调。此外,与绿叶相比,白化叶片中顺式乌头酸、柠檬酸和α-酮戊二酸的浓度都有所增加,苹果酸含量有所降低。这些结果表明大部分的磷酸烯醇式丙酮酸都转移到三羧酸循环中,导致磷酸烯醇式丙酮酸在白化期间较少地转移到莽草酸途径中,并降低了叶片中多酚的代谢。而参与糖酵解和三羧酸循环中相关的的表达水平在白化叶片中均下调,其中包括二氢硫辛基赖氨酸残基乙酰转移酶、二氢硫酰赖氨酸残基琥珀酰基转移酶和二氢硫辛基脱氢酶基因等[2]。在光敏型白化茶树御金香的研究结果发现类黄酮合成途径的上游基因在白化叶片中的表达含量显著提高,其中包括查尔酮合成酶、黄酮类化合物3-羟化酶和查耳酮异构酶以及类黄酮3′,5′-羟化酶。然而在类黄酮合成途径的下游基因中发现无色花色素还原酶、花青素合成酶和花青素还原酶表达含量在白化叶片低于正常叶片。这些研究结果显示在白化茶树御金香中黄酮类通路基因的转录变化有利于槲皮素的生物合成,而不是儿茶素的生物合成[4]。
2.引言
2.1 研究的目的及意义
茶树是一种于世界广泛种植且具有经济效益的作物。茶树也是一种叶用植物,现发现茶树叶片存在着正常叶色(绿色)、黄色、白色、紫色和复色等颜色表型。人们对特定颜色(白色、黄色和紫色等)的茶树进行了广泛的研究,发现叶色不同的茶树其内含成分存在一定的差异,并且具有特定叶色的茶叶的特征化学成分与茶叶质量密切相关。白化茶树是一种叶绿素生物合成或降解受阻、叶绿体发育异常且叶色呈现白色或者黄色等的茶树种质资源。根据目前的研究表明,白化茶树可以分为温敏型白化茶树、光敏型白化茶树和生态不敏感型白化茶树。白化茶树中的白化叶片所制成的绿茶产品具备着高氨基酸,低多酚等优良特点,并且经济效益高。
在黄山群体种的茶园中发现花白茶树单株,变异的叶片中部分组织呈现白色,叶脉呈现绿色。该白化规律不同于温敏型白化茶树的安吉白茶,具有白化期和返绿期。也不同于黄金芽等光敏型白化茶树会随着光照强度变化而改变叶色,经过修剪后发现该花白性状依然存在。此茶树性状特殊,作为观赏性植株或者成为新产品具有很大的潜力。本研究以花白茶树中白化叶片与正常叶片作为研究材料,观察探究其生理生化变化,结合转录组和蛋白组获取差异表达基因和差异蛋白,进一步分析其变异机理,为改善茶树风味品质,为深入了解花白茶树白化机理提供了依据,对理解茶树白化的机理有一定的参考价值。
2.2 研究的内容
(1)花白茶树的内含成分与组织结构差异研究
选取花白茶树的白化叶片和正常叶片为材料,通过石蜡切片法对两者的组织结构进行观察;利用透射电子显微镜观察进行分析研究白化叶片和正常叶片的叶绿体结构差异。测定变异叶片与正常叶片中游离氨基酸、儿茶素及叶绿素关键的生物化学成分。从以上几个方面进行探讨花白茶树的不同叶色组织在生理生化上的差异。
(2)花白茶树的转录组和蛋白组学分析研究
采取花白茶树的白化叶片和正常叶片,利用转录组(RNA-seq)测序和蛋白组学技术,分别与参考基因组进行比对并进行表达差异分析,从而获取花白茶树中白化叶片和正常叶片差异表达基因与差异蛋白。通过功能注释和KEGG富集分析花白茶树不同组织在分子上变化的差异,探讨花白茶树在分子层面上的变化差异。
(4)研究花白茶树的Clp蛋白酶复合体相关基因的表达
通过与拟南芥的基因组和蛋白数据库进行同源序列比对与分析,并从茶树(舒茶早)基因组数据库获得茶树的相关Clp蛋白酶复合体基因序列。利用荧光定量PCR进行测定Clp蛋白酶复合体相关基因的表达,探究花白茶树不同组织结构中Clp蛋白酶复合体相关基因的表达差异。
(5)克隆相关Clp蛋白酶复合体基因及序列分析
选取花白茶树的白化叶片与正常叶片为材料,进行克隆CsClpP5、CsClpP6、CsClpR2基因。克隆所得到的基因全长序列将使用生物信息学进行序列比对和分析。
2.3 技术路线


图2 1技术路线
Fig.2-1 Technical route
3.材料和方法
3.1植物材料
本实验材料采自于安徽省黄山市芙耿城镇辅村茶园中花白茶树单株,该花白茶树不同于温敏型和光敏型白化茶树会随着温度和光照强度变化而改变叶色。采摘花白茶树中芽下第二叶的花白叶片与正常叶色的叶片,分别标记于DL和CDL。花白叶片中的大部分组织产生白化现象,而叶脉呈现绿色。每组样品三个生物学重复。

图3-1 花白茶树的表型
Fig.3-1 The phenotype of albino tea plant
3.2 仪器与试剂
3.2.1主要试剂
XTEDIA(天地)公司:分析试剂色谱级的乙腈、甲醇和乙酸;Wako(和光纯药株)公司:氨基酸混合标准溶液(B型)。
标准品:上海源叶生物科技有限公司:L-茶氨酸(纯度 > 98%)、L-谷氨酸购自;Sigma公司:儿茶素类和咖啡碱标准品。
Watsons公司:蒸馏水;TIANGEN(天根生化)公司:RNAprep pure Plant Kit;北京艾德莱生物科技有限公司(Aidlab Biotechnologies Co.,Ltd):新型植物基因组DNA快速提取试剂盒。
Takara公司:PrimeScript II 1st Strand cDNA Synthesis Kit、PrimeScript™ RT Master Mix Perfect Real Time、TB Green™ Premix Ex Taq™ II(Tli RNaseH Plus)、rTaq酶、DNA Marker(2 Kb);TOYOBO(东洋纺,上海)公司:KOD-FX-Neo高保真酶;Axygen(爱进思,X)公司:DNA凝胶回收试剂盒和质粒提取试剂盒;TRANS公司: pEASY-Blunt Zero克隆载体、Trans1-T1感受态细胞。
3.2.2 主要仪器
奥豪斯仪器上海有限公司:电子天平(AR124CN);
昆山市超声仪器有限公司:数控超声波清洗器(KQ-500DE);
贝克曼公司:高速冷冻离心机;
Hitachi(日本)公司:L-8900全自动氨基酸分析仪,柱长为60 mm,内径4.6 mm以及3 μm的填充颗粒的离子交换树脂柱和UV-5100分光光度计;
X安捷伦(Agilent)公司:高效液相色谱仪(Agilent 1260);
三洋(SANYO,日本)公司:超低温冰箱(FormaTM 8800)、高压灭菌锅;
Bio-Rad(伯乐)公司:微量核酸定量仪、PCR仪、凝胶成像系统、荧光定量PCR仪(Bio-Rad CFX96);
六一仪器厂(北京):电泳仪(DYY-6C);
一恒公司(上海):电热恒温水槽(DK-8D);
天津欧诺仪器仪表有限公司:立式恒温摇床(HNY-2102C);
3.3 茶树内含成分与组织结构的分析
3.3.1叶绿素测定方法
称取茶树鲜叶样品的花白叶片和正常叶色的叶片各100 mg(精确到0.01 mg),将样品剪成细条后立即放入含有10 mL 80%乙醇溶液中的棕色小容量瓶中,常温放置12 h,直至于叶片变白。利用UV-5100分光光度计(Hitachi, Japan)测定其在645 nm和663 nm下的吸收值[73]。在每个样品都要做生物学重复三次,并且每次的生物学重复都要进行三次的技术性重复。
Chla (mg/L)=0.0127D663-0.00269D645
Chlb (mg/L)= 0.0229D645-0.00468D663
Total-Chl(mg/L) = 20.2D645+ 8.02D663
3.3.2游离氨基酸的检测方法
将冻干茶样磨成细粉末,称取20 mg茶样和加入1mL4%的磺基水杨酸于5 mL的离心管,随后在超声波超声仪中超声浸提30 min。在室温条件下,在以每分钟转速12000 的条件下将混合样品进行30 min离心后收集上层清液。重复上述操作再次进行浸提沉淀的部分,而后将两次提取的上清液合并并进行混匀,添加提取液定容至2 mL。合并后的2mL提取液需要通过0.22 μm的水相膜过滤才能装入棕色进样小瓶,进行低温避光保存,备用待测。
利用全自动氨基酸分析仪和4.6×60 mm, 3 μm离子交换树脂柱,标准品L-茶氨酸、L-谷氨酸和氨基酸混合标准溶液(B型)进行游离氨基酸的测定,每个样品测三个生物学重复。
3.3.3茶叶样品中儿茶素类和咖啡碱含量测定方法
将60 mg 冻干样品置于洁净研钵中,并加入1 mL含有80%甲醇和1%的乙酸提取液,充分研磨至匀浆。用2~3 mL 提取液将研磨成的匀浆洗涤到5 mL的离心管中,以每分钟13000 r的转速进行离心并且浸提10 min。收集上清液,用提取液将其定容至4 mL。定容后的4mL提取液需要经过0.22 μm水相膜过滤后装入棕色进样小瓶,低温避光保存待测备用。利用Agilent 1260 高效液相色谱仪和使用反相C18色谱柱(Phenomenex,250×4.6 mm, 5 μm)进行检测分析儿茶素类与咖啡碱在花白茶树中正常对照和花白叶片的含量,其检测方法和计算其含量的方法详见Lu 等[66].
3.3.4茶树叶片的组织结构与超微结构观察
对花白叶片及正常叶片进行组织形态分析需要采用石蜡切片的方法,其中的步骤包含采取样品、样品的固定、洗涤和脱水、透明、浸蜡、包埋、切片与粘片、脱蜡、染色、脱水、透明和封片等将样品制作成切片。样品的每张切片需要利用图像处理软件(Image-Pro Plus 6.0)分别对这随机挑选出的5处视野进行测量5项的形态学特征,其中包括花白叶片和正常对照叶片的厚度、表皮厚度、角质膜厚度和栅栏组织厚度以及海绵组织厚度,其中叶片结构疏松度的计算方法为:(海绵厚度/叶片厚度)×100%。
为了观察叶绿体的超微结构,取茶树样品的叶片,避开主脉,利用刀片切取1平方厘米大小的叶片,用注射器将其浸泡在2.5%戊二醛溶液内。具体方法参见Li等[71]。超薄切片按Daddow等[74]方法进行染色,用HT-7700透射电子显微镜(Hitachi)观察。
3.4转录组分析
3.4.1总RNA的提取和质量检测
采摘花白叶片DL和正常叶片CDL(每个样品三个生物学重复),立即用液氮冷冻的状态下进行研磨。提取茶叶样品总RNA的方法是采用改良后的CTAB方法[75]。利用琼脂糖凝胶电泳,GeneQuant pro RNA/DNA calculator进行检测RNA的浓度和纯度。检测样品的RIN值(RNA的完整值)、28S/18S(RNA完整性的指标)和片段大小是使用Agilent 2100 Bioanalyzer仪器进行的。
3.4.2文库的构建及高通量测序
首先将采用Oligo的磁珠来进行富集茶叶样品的mRNA,随后将得到茶叶样品mRNA在高温和加入打断试剂适量的条件下中进行片断化处理。然后再以打断后的mRNA作为模板合成第一条链cDNA,而后采用二链合成的反应体系来进行合成二链cDNA。使用纯化试剂盒进行回收合成后的双链cDNA,并进行粘性末端修复,在cDNA的3’末端处添加上 A碱基并连接上接头,随后使用PCR方法进行扩增出已选择得到片段大小。现将构建好的文库再次利用Agilent 2100 Bioanalyzer仪器和ABI Step One Plus Real-Time PCR System进行质检,合格后再将对样品进行高通量测序。
3.4.3数据分析以及差异表达基因的筛选
将开始所测序得到的数据被称之为原始数据(raw reads)。首先,我们需要将从原始数据中过滤掉质量不合格和接头有污染以及未知碱基(未能检测)含量过高的片段,并且我们将过滤后的数据称为clean reads。然后将过滤后的数据比对到茶树(舒茶早)参考基因组上,之后再将进行新转录本的预测。在得到新转录本之后挑选出具有蛋白编码潜力的新转录本加入到参考基因序列中构成一个完整的参考序列,然后计算在各个样品中基因的表达水平。根据样品间的比较检测出两者之间差异表达的基因,随后并对差异表达的基因做一系列的生信分析,其中包括GO(Gene ontology)功能分析、Pathway功能分析和转录因子预测等。完整的分析流程图见图3-2。

图3-2 RNA-Seq的信息分析流程图
Fig.3-2 Flow chart of transcriptome resequencing information analysis
使用HISAT软件[76]进行与茶树(舒茶早)基因组进行比对,并使用 Bowtie2[77]软件将过滤后的数据比对到参考序列以统计基因比对率,之后再使用 RSEM[78]计算基因和转录本的表达水平,并采用FPKM方法来进行度量基因的表达水平[79]。使用R软件(version 2.15)里的cor函数计算每样品基因之间的Pearson相关系数并用hclust函数进行层次聚类分析[80]。差异表达基因分析是采用DEseq2 算法,并采用Fold Change >= 2.00 和 Adjusted Pvalue <= 0.05的标准进行筛选出差异表达基因。
根据样品间的差异表达基因,进行通过GO功能分析、KEGG Pathway功能富集分析以及转录因子预测等手段,分析样品(差异表达基因)即在分子层面上、生物功能和代谢途径等的变化。使用Blast2GO(v.2.3.5)对样品间差异基因进行GO功能注释,根据GO注释结果及GO数据库进行分类[81],将其差异表达基因进行功能分类。根据对样品间的差异表达基因的KEGG注释结果以及官方分类,将差异表达基因进行生物通路分类。利用phyper函数对KEGG分类和GO分析结果进行富集分析,并采用显著值(p value)来对富集分析的结果进行校正FDR。将采用getorf软件(EMBOSS:6.5.7.0)并通过检测每一个差异表达基因的开放阅读框来进行预测样品间的差异基因转录因子相关的编码能力。利用转录因子蛋白结构域数据库(PlntfDB)和使用hmmsearch(v. 3.0)软件将差异基因的开放阅读框来进行比对和通过对转录因子家族的特征来明确差异表达基因的编码能力[82]。
3.5蛋白组分析
3.5.1样品的制备方法
采用丙酮沉淀和SDT 裂解法[83]进行处理茶树叶片中的蛋白。
1. 在液氮的条件下,放置适量的茶树样品(CDL;DL)于研钵进行研磨成细粉状态,随后加入5倍丙酮的体积进行混匀样品,混匀后并置在-20℃的冰箱中进行沉淀静置4个小时以上的时间。
2. 静置后的样品在4℃ 的离心机上以6000 g的转速进行离心时间长达40分钟,舍弃上层澄清液体,随后留下的沉淀加入已经在4℃中提前预冷过的丙酮,进行冲洗沉淀三次,后在通风橱中进行吹干沉淀的液体部分。
3. 利用Votex 重悬沉淀的方法在干燥的沉淀粉末(20mg)和SDT裂解液(30倍体积m/v)的混样中进行茶叶样品的细胞分离,而后进行5分钟的沸水浴。
4. 将沸水浴后的混合样品在80 W的超声下进行破碎,期间超声破碎时间为10秒而间隔时间长达15秒,而这超声破碎的工作循环需要进行10次,随后进行时间长达15 min沸水浴。
5. 在以转速14000 g的条件下进行离心样品40分钟,利用0.22 µm滤膜进行过滤上层清液。
6. 样品中蛋白质定量是利用BCA法所进行。随后进行分装样品并在-80℃的超低温的冰箱中进行保存。利用SDS-PAGE 电泳进行蛋白样品纯度的检测。
采用FASP酶解法[84]进行酶切消化成多肽片段。从消化过后的肽段取出100 μg,之后按照说明书(Thermo公司)[85]进行TMT标记。利用分级柱(High pH RP-spin)对用TMT标记的肽段进行分级。分级过后的肽段浓度和质量检测需要用OD280仪器。
利用高效液相色谱分离法进行分离样品中的蛋白。缓冲液:0.1%甲酸水溶液(A液),0.1%甲酸乙腈(84%)水溶液(B液)。用A液相(95%)进行平衡色谱柱,流速为300 nL/min。其中缓冲液B液相梯度变化为 0 min-50 min,0%-35%;50 min-55 min,35%-100%;55 min-60 min,100%。
3.5.2差异蛋白筛选以及相关分析
差异表达蛋白的筛选是按照样品间的倍数变化超过1.1倍(P value <0.05)。与转录组分析方法类似,同样进行使用Blast2Go(https://www.blast2go.com/)[86]软件对样品间差异表达的蛋白进行GO功能注释,随后再利用 KAAS (KEGG Automatic Annotation Server) 软件对样品间的显著性差异表达的蛋白质进行 KEGG 通路注释。差异蛋白在KEGG和GO上的富集度的显著水平是需要通过 Fisher’s Exact Test进行精确检验。差异蛋白在KEGG和GO上的富集度就可以在一定程度表明样品在蛋白水平的变化程度。
3.6引物设计和荧光定量PCR
采用qRT-PCR(Quantitative real-time polymerase chain reaction)方法进行检测花白茶树的Clp蛋白酶复合体相关基因的表达差异。利用Primer5软件设计引物如表3-1,送通用公司进行合成引物。
表3-1花白茶树Clp蛋白酶复合体RT-PCR特异性引物
Table 3-1 Gene-specific primers of Clp complex for qRT-PCR in albino tea
| Gene ID | Forward primer (5’→3’) | Reverse primer (5’→3’) | |
| ClpC1(TEA013518.1) | TAGACTGGTTGGTGAGGCTG | GCAAAACCAAACTGACACTTGA | |
| ClpC2(TEA024181.1) | GTTGGAGGGGGATCTAGTG | CACCAGGCTCTCCGATAAG | |
| ClpP2(TEA008049.1) | CTCTCCCTAATGCAACAATAA | CATTCAATACATCCCACACC | |
| ClpP3(TEA013179.1) | CACCTCCACCCAAAACCA | TCCGAGCCTCTCTCCTCA | |
| ClpP4-1(TEA003125.1) | GCAATGCCCAACACACAGAT | GGAGACATGTAGCGATCCCT | |
| ClpP4-2(TEA024098.1) | CTCACTTGCTTCTCACCAATT | AGAACACAATCCTCTCCTTCA | |
| ClpP5(TEA023304.1) | CTTCCCAAACCAAAACCC | CTGAAAACCGACCCGAGT | |
| ClpP6(TEA022442.1) | CTTGAGTTATGGTGCAATAGAA | CCTGTGAGTTTATTGGCTGT | |
| ClpR1(TEA004864.1) | TCGAAGCCTCTTGACCATGT | GACACTTTCAGACTGTCGGC | |
| ClpR2(TEA025416.1) | TCGCTTTGCAATGCCTCTTT | CGCTTCATCCGGCTTAAGTC | |
| ClpR3(TEA025898.1) | TGAATCTCCCACGCTCAT | CCTCACTCTCCATCCCAA | |
| ClpR4(TEA004336.1) | CCCCCTATTTTTACACTCTG | TATGCTTTGAATAGAGTTTAACC | |
使用Prime-Script™ RT Master Mix perfect Real Time试剂盒,将提取的样品总RNA进行定量反转录,选择的内参基因为Csactin-F(5’-3’:GCCATCTTGATTGGAATGG)以及 Csactin-R (5’-3’:GGTGCCACAACCTTGATCTT)。利用TB Green™ Premix Ex Taq™ II Tli RNaseH Plus的荧光定量试剂盒并在CFX96 Realtime-PCR检测系统(Bio-Rad, Hercules, USA)对候选基因进行qRT-PCR实验。
反应体系如下:
| Premix Ex Taq™ II | 12.5 μL |
| cDNA模板 | 2.0 μL |
| Forward primer | 1.0 μL |
| Reverse primer | 1.0 μL |
| ddH2O | 8.5 μL |
| Total | 25 μL |
3.7 Clp相关基因克隆及序列分析
3.7.1总RNA提取和质量检测
同上步骤3.4.1的进行提取花白叶片与正常叶片的总RNA,并进行质量检测。
3.7.2 cDNA的合成
按照TaKaRa的反转录试剂盒的说明书进行合成花白叶片与正常对照的cDNA。进行反转录实验的过程中都需要在4℃的条件下完成,反转录成功后的cDNA需要在-80℃的条件下进行长期的保存。
3.7.3 PCR扩增
参考茶树的基因组数据(舒茶早),从中分析得到ClpP5、ClpP6、ClpR2基因序列。所得的ClpP5、ClpP6、ClpR2参考基因序列需要在NCBI数据库中进行比对,结果发现其基因序列中都具有完整的开放阅读框。故利用Primer5软件设计相关基因的全长引物(表3-2)。
表3-2花白茶树Clp复合体克隆特异性引物
Table 3-2 Gene-specific primers of Clp complex in albino tea
| Gene ID | Forward primer (5’→3’) | Reverse primer (5’→3’) |
| ClpP5(TEA023304.1) | ATGGCGCATTCGTGC | TTACTGATCTGCACTCGCTG |
| ClpP6(TEA022442.1) | ATGAAAAAACTGTTTGCGCAAG | TTAGTATTCTGTTTCCAAGACCCCA |
| ClpR2(TEA025416.1) | ATGGGGAGTCTATTCGCATG | CTAACCAAGACCTGTTCCTG |
目的基因的扩增反应体系:
| 2×PCR buffer | 25 µL |
| 2 mM dNTPs | 10 µL |
| Forward Primer(10 µM) | 1.5 µL |
| Reverse Primer(10 µM) | 1.5 µL |
| KOD Fx Neo | 1 µL |
| cDNA | 1 µL |
| ddH2O | 10 µL |
| Total | 50 µL |
PCR扩增全长基因的反应条件是在94°C的条件下进行预变性5分钟,随后按照98°C的条件下PCR仪运行30秒,而后在60°C的条件下运行30秒,之后在68°C的条件下运行2 min,此过程需要循环35次。随后有需要68°C的条件下再次进行延伸10 min,在4°C的冰箱中进行保存PCR扩增出的产物。PCR扩增的克隆产物需要用1.2%的琼脂糖凝胶进行分析,切胶出扩增出的目标产物凝胶。而后再按照胶回收的指示说明书进行回收胶产物,记录回收后产物的核酸浓度,备用状态的时候需放置于-20°C的低温冰箱中保存。将纯化后的目的基因导入克隆载体(Peasy-blunt-zero),根据转化感受态细胞的说明书指示转化至Trans1-T1感受态细胞中,转化后的菌液需要放置在37°C条件下的振荡培养箱中进行摇晃时间1小时。从略显浑浊的菌液吸取100 µL进行涂布在含有100 µg/mL Amp(抗生素)的LB琼脂板上,并需要在37°C条件下的恒温培养箱中进行培养长达12小时。通过划线检测出阳性克隆的菌斑点,将检测出阳性单克隆进行随机挑选送至通用公司测序。
3.7.4 相关基因的生物学分析
获得的目的基因全长序列在NCBI网站上进行同源性分析,并利用生物学软件DNAMAN和BioXM 2.6进行多序列比对。同时利用在线分析软件TMHMM 2.0和NetPhos 3.1 Server, 对目的蛋白进行预测分析其结构域和磷酸位点。
3.8 CsClpP5寡核苷酸实验
利用网站http://sfold.wadsworth.org/cgi-bin/index.pl设计特定基因的反义寡聚核苷酸引物设计CsClpP5的asODN(antisense oligonucleotide,反义寡核苷酸)序列Anti-ClpP5序列为5’-TGGGAAGAGGAGTTAGGGTT-3’;5’-CCTTGTGATGCAACATTTCA-3’;5’-TTTGCCTTGTGATGCAACAT-3’。选用1.5mL离心管,加入1mL稀释为20uM的反义寡聚核苷酸引物溶液,插入长势良好的一芽二叶离体植株,用透气性封口膜密封管口来进行防止引物溶液挥发,以防实验造成误差;对照为10μM蔗糖溶液;放置好的一芽二叶的离体小枝条需要在室温进行培养长达24小时,取样的时间分别设定在0小时、6小时、12小时和24小时,并且用液氮进行固样,并将样品放置于-80℃的超低温冰箱中进行保存,用作于进行下一步检测分析;该实验不低于3个重复。根据时间点收集的样品进行提取RNA并进行实时荧光定量PCR,检测靶基因表达。
4结果与分析
4.1花白茶树生化成分含量分析
叶绿素能够吸收传递光给植物叶绿体进行光合作用,在植物发育成长的进程中发挥着举足轻重的作用[87]。本实验中针对花白叶片(DL)和正常叶片(CDL)检测叶绿素 a(Chlorophyll a,Chla)和叶绿素 b(Chlorophyll b,Chlb)的含量,如图4-1所示,发现正常叶片的叶绿素a、叶绿素b含量和总叶绿素含量(Chla+ Chlb)均显著高于花白叶片。这表明正常叶片和花白叶片叶色差异较大,进行研究花白茶树的叶色形成和相关的物质代谢变化等分子机制具有一定的价值性。
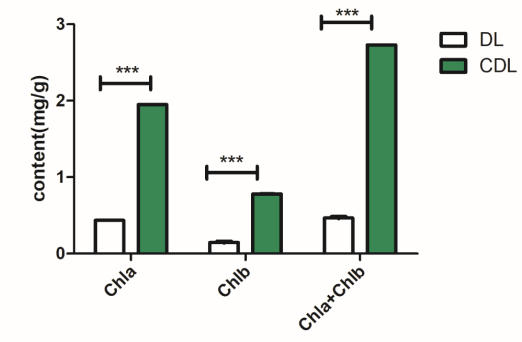
图4-1 花白茶树叶绿素含量分析
Fig.4-1 Analysis of chlorophyll content in albino plant
茶叶中化学成分包括咖啡碱、儿茶素和氨基酸,能够组成其特征性风味,并有一定的保健功能。本实验中针对花白叶片(DL)和正常叶片(CDL)测定了其咖啡碱、儿茶素和游离氨基酸的含量。
如表4-1所示,在检测咖啡碱(Caffeine)含量中,结果表明花白叶片略高于正常叶片。而在检测儿茶素(Catechin,C)、没食子儿茶素(Gallocatechin,GC)、表儿茶素(Epicatechin,EC)、表没食子儿茶素(Epigallocatechin,EGC)、表儿茶素没食子酸酯(Epicatechin gallate,ECG)、表没食子儿茶素没食子酸酯(Epigallocatechin gallate, EGCG)的含量结果中发现正常叶片所含的儿茶素均高于花白叶片。其中,正常叶片的GC含量是花白叶片中的4倍之多。相对于正常叶片,花白叶片中EGC、ECG和C的含量减少了近50%,EGCG的含量减少了33.16%。并且,正常叶片中总儿茶素类的含量是花白叶片的1.5倍。总体来说,花白叶片中儿茶素含量减少,咖啡碱的含量略有增长。
表4-1 花白茶树咖啡碱和儿茶素类含量分析
Table 4-1 Analysis of theine and catechin contents
| Compounds(mg/g DW) | DL | CDL | ||||
| Flavan-3-ols | ||||||
| Catechin (C) | 0.21 | ± | 0.003 | 0.40 | ± | 0.02 |
| Gallocatechin (GC) | 3.71 | ± | 0.17 | 14.61 | ± | 0.52 |
| Epi-catechin (EC) | 4.12 | ± | 0.35 | 5.84 | ± | 0.29 |
| Epi-gallocatechin (EGC) | 1.12 | ± | 0.06 | 2.05 | ± | 0.13 |
| Epi-catechin-3-gallate (ECG) | 7.49 | ± | 0.30 | 13.86 | ± | 0.56 |
| Epi-gallocatechin-3-gallate (EGCG) | 64.40 | ± | 2.40 | 85.76 | ± | 3.50 |
| Total flavan-3-ols | 81.10 | ± | 3.26 | 123.81 | ± | 5.04 |
| Purine alkaloid | ||||||
| Caffeine | 25.64 | ± | 0.55 | 23.69 | ± | 0.85 |
通过对花白茶树中游离氨基酸的检测结果发现花白叶片中检测的氨基酸含量大多高于正常叶片(表4-2)。苏氨酸(Threonine,Thr)、丝氨酸(Serine,Ser)、甘氨酸(Glycine,Gly)、丙氨酸(Alanine,Ala)、色氨酸(Tryptophan,Trp)、赖氨酸(Lysine,Lys)和组氨酸(Histidine,His)在花白茶树中积累的含量在0.01~0.64 mg/g DW。在检测的游离氨基酸中,茶氨酸在花白茶树中积累最多,其次是谷氨酸。其中,天冬氨酸(Aspartic Acid,Asp)在花白叶片中的含量是正常叶片的3倍,在茶叶中含量最丰富的茶氨酸(Theanine,Thea)亦在花白叶片中含量高于正常叶片,含量为18.92 mg/g DW,约为正常叶片(7.05 mg/g DW)的2.6倍。正常叶片中精氨酸(Arginine,Arg)的含量较低,而花白叶片(0.73 mg/g DW)中积累多,是正常叶片的73倍。与之相反,丝氨酸、甘氨酸和谷氨酰胺(Glutamine,Gln)以及丙氨酸在正常叶片中含量积累要比花白叶片的多。丝氨酸的含量在花白叶片中减少了15.38%,甘氨酸则在花白叶片中减少了50%。谷氨酰胺的含量减少了33.02%。结果表明,花白茶树中花白叶片的游离氨基酸含量积累比正常叶片高。
表4-2 花白茶树游离氨基酸含量分析
Table 4-2 Analysis of free amino acid content
| Compounds(mg/g DW) | DL | CDL | ||||
| Amino acids | ||||||
| Aspartic Acid(Asp) | 2.96 | ± | 0.004 | 0.89 | ± | 0.16 |
| Threonine(Thr) | 0.17 | ± | 0.002 | 0.13 | ± | 0.02 |
| Serine(Ser) | 0.39 | ± | 0.001 | 0.45 | ± | 0.07 |
| Glutamic Acid(Glu) | 4.84 | ± | 0.04 | 3.29 | ± | 0.56 |
| Glutamine(Gln) | 1.09 | ± | 0.01 | 1.45 | ± | 0.23 |
| Theanine(Thea) | 18.92 | ± | 0.18 | 7.05 | ± | 1.06 |
| Glycine(Gly) | 0.05 | ± | 0.001 | 0.10 | ± | 0.01 |
| Alanine(Ala) | 0.16 | ± | 0.001 | 0.17 | ± | 0.02 |
| Tryptophan(Trp) | 0.64 | ± | 0.002 | 0.54 | ± | 0.09 |
| Lysine(Lys) | 0.05 | ± | 0.002 | 0.01 | ± | 0.003 |
| Histidine(His) | 0.04 | ± | 0.001 | 0.02 | ± | 0.001 |
| Arginine(Arg) | 0.73 | ± | 0.01 | 0.01 | ± | 0.002 |
| Total free amino acid | 31.10 | ± | 0.22 | 14.88 | ± | 2.35 |
4.2 花白茶树生理变化分析
4.2.1花白茶树组织结构分析
花白茶树组织结构分析是使用石蜡切片进行研究的。通过花白叶片和正常叶片的石蜡切片图(图4-2)可看出其组织结构有着明显的差别。花白茶叶中的栅栏组织细胞排列较疏松,栅栏组织的厚度也略低于正常叶片(表4-3),但无显著性差别。海绵组织亦如此,但测量其两者的厚度没显著性的差别。测量表皮厚度,角质膜厚度,叶片厚度的结果表明,花白叶片中厚度(184.92 μm)显著低于正常叶片(196.58 μm)。与此同时,计算白化叶片的叶片结构疏松度(RL)也是显著低于正常叶片。

图4-2花白叶片(A)和正常叶片(B)的显微结构图
Fig.4-2 The microstructure of the abino leaf (A) and the normal leaf (B)
表4-3 花白茶树组织结构分析
Table 4-3 Analysis of tissue structure of albino tea plant
| 花白叶片 | 正常叶片 | |||||
| 表皮厚度(μm) | 11.86 | ± | 0.91* | 16.98 | ± | 1.06 |
| 角质膜厚度(μm) | 1.72 | ± | 0.29* | 2.38 | ± | 0.38 |
| 栅栏组织厚度(μm) | 29.88 | ± | 3.92 | 33.06 | ± | 2.52 |
| 海绵组织厚度(μm) | 125.54 | ± | 3.92 | 125.96 | ± | 3.55 |
| 叶片厚度(μm) | 184.92 | ± | 4.56* | 196.58 | ± | 2.06 |
| 叶片组织结构疏松度 (RL) | 67.89 | ± | 1.32* | 64.07 | ± | 1.24 |
4.2.2 花白茶树叶片超微结构分析
采用透射电镜进行观察研究花白茶树的叶片中叶绿体超微结构的变化。如图4-3A所示,正常叶片中的叶绿体结构正常,存在着明显的基质类囊体膜(Gr)和相应的片层结构。与此同时还发现叶绿体(Ch)是贴细胞壁(CW)生长,淀粉粒(SG)清晰可见。相比较于正常叶片,观察结果发现花白叶片的叶肉细胞中叶绿体发育异常,叶绿体结构存在空腔并且没有明显的基质类囊体(图4-3B)。
图4-3正常叶片(A)和花白叶片(B)的叶绿体超微结构
Fig.4-3 Chloroplast ultrastructure of normal leaf (A) and albino leaf (B)
4.3 花白茶树转录组分析
4.3.1 测序数据的质量评估与参考基因组比对
本研究使用Illumina HiSeq平台进行检测了6个样品,如表4-4所示,每个样品平均产出6.71 Gb数据。其中检测的结果显示测序质量指标(Q20值和Q30)均达到93%以上,表明该次所测序数据质量高并且可信度高,数据可用于后续分析实验。此外,从表4-4中还发现花白茶树(正常叶片,CDL和花白叶片,DL)比对茶树基因组的平均比对率为80.40%。
表4-4 转录组测序质量评估
Table 4-4 quality assessment of transcriptome sequencing
| Sample | Total
Raw Reads(Mb) | Total
Clean Reads(Mb) | Total
Clean Bases(Gb) | Clean Reads Q20(%) | Clean Reads Q30(%) | Clean Reads Ratio(%) |
| CDL_1 | 53.9 | 44.76 | 6.71 | 98.12 | 94.06 | 83.06 |
| CDL_2 | 52.26 | 44.38 | 6.66 | 98.2 | 94.47 | 84.91 |
| CDL_3 | 53.9 | 44.26 | 6.64 | 98.31 | 94.82 | 82.12 |
| DL_1 | 52.26 | 44.9 | 6.74 | 98.04 | 93.77 | 85.92 |
| DL_2 | 52.26 | 44.9 | 6.73 | 98.03 | 93.75 | 85.91 |
| DL_3 | 50.63 | 44.77 | 6.71 | 98.54 | 95.28 | 88.42 |
4.3.2差异表达基因GO功能分析
采用GO的定义方法对花白叶片和正常对照中的差异表达基因进行功能注释。在CDL-VS-DL中,总共获得了3431个DEGs;在这些序列中,1619个上调,1812个下调。在生物过程类中,“代谢过程”、“单个生物过程”和“细胞过程”是最富集的GO分析,而在细胞成分类中,“细胞”、“细胞部分”和“膜”在花白茶树中的变化最显著。相应的在分子功能类中,从结果中可以看出“催化活性”和“结合”在变化最明显(图4-4)。

图4-4 花白茶树中差异基因GO富集分析
Fig.4-4 GO enrichment analysis of DEGs in CDL-VS-DL
4.3.3 差异表达基因在KEGG途径中的富集分析
KEGG Pathway数据库是KEGG数据库的子数据库,它整合了不同物种中的代谢通路及相关的基因信息。由图4-5所示,在CDL-VS-DL中,发现代谢途径的差异表达基因的最多且差异较为显著,其次为次级代谢差异显著。从差异基因KEGG富集分析的结果发现,有关于碳代谢中的黄酮类生物合成、芪类,二苯基庚酮和姜酚生物合成、淀粉和蔗糖代谢、戊糖,葡萄糖醛酸转换、亚麻酸代谢途径、柠檬烯,蒎烯降解、类倍半萜烯和三萜生物合成、亚麻酸代谢、果糖和甘露糖代谢和花青素生物合成受到显著的富集,表明在花白茶树中的碳代谢受到了影响。同样的也发现差异基因在关于氮代谢的色氨酸代谢、咖啡碱代谢和氰基氨基酸代谢进行了显著的富集。关于初级代谢的核糖体和内质网蛋白加工途径中的差异表达基因变化明显。
图4-5 花白茶树中差异基因KEGG富集分析
Fig.4-5 KEGG enrichment analysis of DEGs in CDL-VS-DL
4.3.4 茶叶关键化合物的代谢途径中基因表达分析
茶氨酸是白化茶叶中积累的一种关键风味化合物[71, 88]。茶氨酸生物合成的编码基因(图4-6A),包括茶氨酸合成酶(TS)、谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)、硝酸还原酶(NR)、亚硝酸还原酶(Nir)、谷氨酸脱氢酶(GDH)、丙氨酸转氨酶(ALT)和丙氨酸脱羧酶(ADC)。与正常绿叶相比,花白叶片中茶氨酸合成酶(TS)的表达没有明显变化,Nir的表达略微下调。相比之下,其余的谷氨酰胺合成酶(GS)、谷氨酸合成酶(GOGAT)、硝酸还原酶(NR)、谷氨酸脱氢酶(GDH)、丙氨酸转氨酶(ALT)和丙氨酸脱羧酶(ADC)的表达均有上调,这可能会提高白化茶叶中的茶氨酸含量[89]。
类黄酮化合物是决定茶叶风味的特征成分[4]。为了更好地了解花白茶叶片中类黄酮的积累,我们对类黄酮生物合成途径中的关键基因进行了研究(图4-6B)。与正常叶片相比,花白叶片的二氢黄酮醇4-还原酶(DFR)基因表达略微上调。花白叶片中的儿茶素生物合成途径上游基因呈现出低表达的状态。这些基因包括苯丙氨酸氨解酶(PAL)、4-香豆酰辅酶A连接酶(4Cl)、肉桂酸4-羟化酶(C4H)和黄酮类化合物3-羟化酶(F3H)以及查尔酮合成酶(CHS)。在花白叶片中,类黄酮生物合成途径中的下游基因花青素合成酶(ANS)和花青素还原酶(ANR)的基因表达也低于正常叶片。这表明与类黄酮合成相关的基因低表达可能与花白叶片中儿茶素含量的降低有关。

图4-6 花白茶树中茶氨酸生物合成途径(A)和黄烷-3-醇生物合成途径(B)
Fig.4-6 Expression levels of key genes in the theanine biosynthetic
pathway (A) and the flavan-3-ol biosynthetic pathway (B).
叶绿素参与植物的光合作用,同时也在植物生长发育和叶色变化中起到重要的作用。在CDL_VS_DL中,发现3条差异表达基因参与叶绿素合成途径。如图4-7A所示,花白叶片中叶绿素a加氧酶(CAO)、香叶酰二磷酸还原酶(CHLP)和原叶绿素氧化还原酶(POR)的表达含量均显著低于正常叶片。这表明与叶绿素合成相关的差异基因低表达可能与花白叶片中叶绿素含量的降低有关。
在光合作用中,有9条差异表达基因(图4-7B)。光系统II的捕光复合体(LHCB1和LHCB2),光系统I的捕光复合体(LHCA2)和铁氧化蛋白(PetF)在花白叶片表达含量均低于正常叶片。而其余的光系统II CP47叶绿素脱辅蛋白(PsbB)和ATP合酶(ATPD和ATPG)在花白叶片表达含量显著高于正常叶片。

图4-7 花白茶树中叶绿素合成途径(A)和光合作用(B)
Fig.4-7 Chlorophyll synthesis pathway (A) and photosynthesis (B) in albino tea plant
4.3.5 转录因子的分析
转录因子可以在转录水平上进行调控基因的表达,可通过功能域等其他方式来激活或者抑制基因的表达[90]。在CDL-VS-DL中,鉴定出了40个转录因子家族(图4-8)。相对于正常叶片来说,在白化叶片中发现有179个上调的转录因子,160个下调。花白叶片中上调的转录因子主要包括了有关于AP2-EREBP家族29个,MYB家族为18个,WRKY家族是17个,GRAS家族13个,bHLH家族11个,MYB-related家族11个和TCP家族10个。相对的下调转录因子主要集中于AP2-EREBP(18个),bHLH(17), MYB(16个)与MYB-related(16个)。

图4-8 花白茶树中转录因子家族的鉴定
Fig.4-8 Identification and categorization of TF families in CDL-VS-DL in albino tea plant
4.4 花白茶树蛋白组学分析
4.4.1 差异表达蛋白的筛选结果
从花白叶片和正常叶片中总共鉴定出6688个蛋白,通过标准进行筛选出差异表达蛋白,比较组DL_VS_CDL的差异表达蛋白有95个,其中发现在花白叶片中53个上调和42个下调的差异表达蛋白。
4.4.2 差异蛋白关于GO功能的富集分析
如图4-9所示,前20个显著富集GO功能的结果表明在比较组DL_vs_CDL 中的氧化磷酸化(oxidative phosphorylation),磷酸化作用(phosphorylation),含嘌呤化合物代谢过程(purine-containing compound metabolic process),adenine salvage(腺嘌呤回补)和碳水化合物衍生物代谢过程(carbohydrate derivative metabolic process)等重要生物学过程,氧化还原酶活性 (oxidoreductase activity), 作用于硫基供体的活性(acting on a sulfur group of donors),查尔酮异构酶活性(chalcone isomerase activity),3-氧酰基-[酰基载体蛋白]合成酶活性等分子功能(3-oxoacyl-[acyl-carrier-protein] synthase activity),嘌呤磷酸核糖转移酶活性(purine phosphoribosyltransferase activity)和腺嘌呤磷酸核糖转移酶活性(adenine phosphoribosyltransferase activity)等分子功能,发生了显著性变化。

图4-9 花白茶树中差异蛋白GO富集分析
Fig.4-9 Enrichment analysis of different expressed protein GO in albino tea tree
4.4.3 差异表达蛋白KEGG通路富集分析
如图4-10所示,采用Fisher的检测方法让其对对其花白叶片与正常对照的差异表达蛋白进行KEGG 通路的富集分析。之后在差异蛋白富集的结果发现氧化磷酸化、类黄酮生物途径和苯丙素生物合成的这三个重要通路在花白叶片与正常对照中发生了显著变化。同时确定了许多的途径存在差异,包括光合作用,嘌呤代谢等代谢途径(表4-5)。其中,差异倍数最大的差异蛋白是与病原相关的PR蛋白(Pathogenesis-related protein PR-4-like),花白叶片中PR蛋白的含量是正常叶片的1.4倍。除此之外,还发现花白叶片在有关于植物病原抵御中的几丁质触发子受体激酶(Chitin elicitor receptor kinase)蛋白含量也高于正常叶片。表明花白茶树中花白叶片可能处于逆境中。有关于合成氨基酸代谢中发现参与甘氨酸、丝氨酸和苏氨酸的代谢途径的D-3-磷酸甘油酸脱氢酶(D-3-phosphoglycerate-dehydrogenase, PGDH)在花白茶树中有所积累,蛋白含量是正常叶片的1.16倍。有关于碳代谢中的类黄酮生物合成途径、苯丙素生物合成途径、甘油磷脂代谢途径以及脂肪酸生物合成途径和倍半萜和三萜生物合成途径中的相关差异蛋白在花白叶片中均处于下调。而有关于甘油酯类的代谢中蛋白在花白叶片中处于上调,是正常叶片的1.13倍。另外,还发现有关于初级代谢中的核糖体生物合成、RNA转运、细胞的胞吞作用和内质网加工蛋白等活动在花白茶树中受到影响,其相关的蛋白在花白叶片中上调,大约是正常叶片的1.2倍。而相关于过氧化物酶的蛋白在花白叶片中处于下调,是正常叶片的0.9倍。

图4-10 花白茶树中差异蛋白KEGG富集分析
Fig.4-10 Enrichment analysis of different expressed protein GO inalbino tea tree
表4-5 在不同途径中鉴定的差异积累的蛋白质
Table 4-5 Differentially accumulated proteins identified in pathways
| Identity Protein | Protein ID | EC Number | Fold Changes (DL/CDL) |
| Flavonoid biosynthesis | |||
| Chalcone isomerase | TEA033023.1 | EC:5.5.1.6 | 0.84 |
| Anthocyanidin reductase | TEA022960.1 | EC:1.3.1.77 | 0.81 |
| Photosynthesis | |||
| PetF( ferredoxin) | TEA005268.1 | 0.89 | |
| F-type H+-transporting ATPase subunit b | TEA006995.1 | 1.11 | |
| Oxidative phosphorylation | |||
| NADH-ubiquinone oxidoreductase chain 5 | TEA025710.1 | EC:7.1.1.2 | 1.30 |
| NADH dehydrogenase (ubiquinone) FeS protein 6 | TEA009800.1 | 1.25 | |
| Cytochrome c oxidase subunit 1 | TEA030457.1 | EC:1.9.3.1 | 0.64 |
| F-type H+-transporting ATPase subunit b | TEA006995.1 | 1.11 | |
| Purine metabolism | |||
| Adenosine kinase | TEA001315.1 | E2.7.1.20 | 0.91 |
| Pyruvate kinase | TEA010980.1 | EC:2.7.1.40 | 1.29 |
| Adenine phosphoribosyltransferase | TEA017768.1 | EC:2.4.2.7 | 0.86 |
| Glycine, serine and threonine metabolism | |||
| D-3-phosphoglycerate dehydrogenase | TEA014195.1 | EC: 1.1.1.399 | 1.16 |
| Phenylpropanoid biosynthesis | |||
| Peroxidase | TEA026798.1 | EC1.11.1.7 | 0.86 |
| beta-glucosidase | TEA025857.1 | EC:3.2.1.21 | 0.80 |
| Cinnamoyl-CoA reductase | TEA032454.1 | EC:1.2.1.44 | 0.78 |
| Glycerolipid metabolism | |||
| 1,2-diacylglycerol 3-beta-galactosyltransferase | TEA014152.1 | EC:2.4.1.46 | 1.13 |
| Glycerophospholipid metabolism | |||
| Phospholipase C | TEA004903.1 | EC:3.1.4.3 | 0.73 |
| Fatty acid biosynthesis | |||
| 3-oxoacyl-[acyl-carrier-protein] synthase III | TEA021479.1 | EC:2.3.1.180 | 0.81 |
| Acting on a sulfur group of donors | |||
| Thioredoxin 1 | TEA014772.1 | 0.86 | |
| Thioredoxin 1 | TEA002799.1 | 0.80 | |
| Sesquiterpenoid and triterpenoid biosynthesis | |||
| Beta-amyrin synthase | TEA026265.1 | EC:5.4.99.39 | 0.79 |
| Ribosome biogenesis in eukaryotes | |||
| U3 small nucleolar RNA-associated protein MPP10 | TEA013154.1 | 1.24 | |
| RNA transport | |||
| Cytoplasmic FMR1 interacting protein | TEA001169.1 | 1.28 | |
| Plant-pathogen interaction | |||
| Chitin elicitor receptor kinase 1 | TEA030161.1 | 1.11 | |
| Defense response | |||
| Pathogenesis-related protein PR-4-like | TEA013238.1 | 1.40 | |
| Endocytosis | |||
| ESCRT-I complex subunit VPS37 | TEA018005.1 | 1.29 | |
| Peroxisome | |||
| Dehydrogenase/reductase SDR family member 4 | TEA013806.1 | EC:1.1.-.- | 0.90 |
| Protein processing in endoplasmic reticulum | |||
| GTP-binding protein SAR1 | TEA024772.1 | EC:3.6.5.- | 1.16 |
4.5 花白茶树的Clp蛋白酶复合体
4.5.1 花白茶树Clp蛋白酶复合体及其实时荧光PCR
根据模式物种拟南芥中Clp蛋白酶复合体相关的基因序列,比对舒茶早茶树基因组,找到了与拟南芥同源的Clp基因。其中包括ClpP亚基(ClpP2、ClpP3、ClpP4-1、ClpP4-2、ClpP5和ClpP6)、4个ClpR亚基(ClpR1到ClpR4),2个ClpC (CIpC1和ClpC2)。
以舒茶早叶片(SCZ)和花白茶树的正常叶片做对照,比较Clp复合体相关基因在花白叶片中表达。如图4-11所示,ClpR2基因表达水平在舒茶早和花白茶树中最高,其次为ClpP5的基因表达,相反的是ClpC-2基因表达含量在舒茶早和花白茶树中最低。其中ClpR2在花白和正常叶片中的表达显著低于舒茶早,而在花白叶片和正常叶片中并无明显差异。类似的,舒茶早中的ClpC-1和ClpR4基因表达水平显著高于正常叶片和花白叶片。ClpP5基因的表达在花白叶片中显著高于正常叶片和舒茶早,而在正常叶片和舒茶早中并无明显差异。ClpP4-2、ClpR3和ClpP6基因的表达趋势与ClpP5的类似,在花白叶片中表达最高。而ClpP2和ClpP3的基因表达含量在舒茶早最高。ClpP4-1基因的表达在舒茶早、正常叶片和花白叶片中并无显著性差异。

图4-11 花白茶树中Clp蛋白酶复合体基因表达分析
Fig.4-11 Gene expression analysis of Clp complex
4.5.2 花白茶树Clp蛋白酶复合体相关基因的克隆
根据研究表明:当拟南芥Clp蛋白酶复合体中的ClpP6和ClpR2基因发生突变,会导致拟南芥植株生长滞缓,叶色黄化[91]。由图4-11可知ClpR2基因在舒茶早和花白茶树的Clp蛋白酶复合体中表达最高,ClpP6基因的表达花白茶树中存在差异。故以花白叶片和正常叶片为模板,进行扩增CsClpP6基因(图4-12A)与CsClpR2基因(图4-12B)。从花白叶片和正常叶片测序拼接得到CsClpP6基因全长序列均为852bp,其CsClpP6基因编码284个氨基酸。同样的,CsClpR2基因全长序列也均为1671bp并且编码557个氨基酸。经过比对花白叶片和正常叶片克隆出的CsClpP6与CsClpR2基因均无变异。

图4-12 PCR琼脂糖凝胶电泳检测图
Fig. 4-12 Agarose gel electrophoresis
玉米中ClpP5基因发生突变,也会使得玉米幼苗发生白化[45],故其以花白叶片和正常叶片为模板,进行扩增CsClpP5基因(图4-13)。拼接结果显示花白叶片中扩增的CsClpP5全长基因序列为909bp,共编码303个氨基酸。然而正常叶片中扩增的CsClpP5基因全长903bp并且编码301个氨基酸(图4-14)。
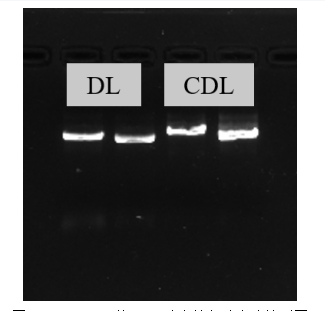
图4-13 CsClpP5的PCR琼脂糖凝胶电泳检测图
Fig. 4-13 Agarose gel electrophoresis of CsClpP5 gene

图4-14 CsClpP5基因编码氨基酸比对图
Fig. 4-14 Map of amino acids encoded byCsClpP5 gene
4.5.3 花白茶树CsClpP5生物信息学分析
利用SignalP-3.0 Server进行预测CsClpP5蛋白质,结果发现CsClpP5蛋白质信号肽S值均小于0.5并且其预测的信号肽曲线表现出不典型状态,故进而推测出CsClpP5蛋白质不具有信号肽结构。CsClpP5蛋白质跨膜结构是通过TMHMMServerV.2.0在线软件进行预测的,发现其跨膜区域存在可能性小,预测CsClpP5蛋白质属于非跨膜蛋白。利用NetPhos 3.1 Server在线软件对正常叶片克隆出的CsClpP5蛋白质进行预测磷酸化位点分析,预测分析结果表明其蛋白质包含着36个磷酸化位点,其中包括着26个丝氨酸磷酸化、6个酪氨酸磷酸化和4个苏氨酸磷酸化。而花白叶片中的CsClpP5蛋白质共预测出37个磷酸化位点,其中存在着丝氨酸磷酸化28个、酪氨酸磷酸化6个和苏氨酸磷酸化3个。说明花白叶片的CsClpP5蛋白质比正常叶片的增加了两个丝氨酸,会改变CsClpP5蛋白质磷酸化位点的变化。
从NCBI上下载相似度比较高的ClpP5蛋白质,其中有拟南芥、番木瓜、蔡麦及毛霉的ClpP5蛋白的同源序列。利用DNAMAN软件进行匹配比对ClpP5蛋白的同源序列,结果如图4-15,保守区域在100到300氨基酸,活性位点为TVCVGLAASMGA。花白叶片中CsClpP5并未突变在活性位点上。
图4-15 CsClpP5与其他物种同源序列的比对图
Fig. 4-15 Multiple alignment of amino acid of CsClpP5 and homologous genes in other species
注:下划线为活性位点
Note: the underline is the active site
4.5.3 花白茶树CsClpP5反义寡核苷酸实验
为了了解茶树CsClpP5对于茶树叶色的影响,设计CsClpP5的asODN序列。让茶树叶片吸收anti-CsClpP5,观察处理组叶色变化,并分别在0小时,6小时,12小时和24小时这4个时间点进行取样,利用实时荧光定量PCR方法进行检测CsClpP5基因在其实验中的表达水平变化。观察叶色结果结果为,长时间的吸收anti-CsClpP5,未能使得茶树叶色发生变化。荧光定量结果表明(图4-16),经过24h并以蔗糖水饲喂的茶树叶片即对照,CsClpP5表达水平差不多。但处理组与对比组相比,发现6h的处理组中CsClpP5表达水平下降,随后有上升的趋势。说明在短时间内,anti-CsClpP5能使得茶树中的CsClpP5表达降低,但未能使得茶树叶色发生变化。

图4-16 CsClpP5反义寡核苷酸的定量表达
Fig. 4-16 Quantitative expression of antisense oligonucleotides of CsClpP5
5.讨论
因具有特定叶色的茶叶的特征化学成分与茶叶质量密切相关[6, 92],因此茶树叶色的多样性在近年来成为了研究热点之一。目前已有报道对安吉白茶、白鸡冠、黄金芽、御金香等茶树的叶色变化也进行了研究。早春,安吉白茶的枝条在低温下会变黄或变白,但气温升高时会恢复绿色[8]。黄金芽[93],御金香[4],和白鸡冠[3]等光敏型白化茶树的叶子会在高强度阳光下变黄,遮荫后又会返绿。类似的现象在拟南芥[22],玉米[23],棉花[24],水稻[25],番茄[26]等植株上也有进行报道。
5.1 花白茶树的生理生化分析
叶绿体是植物进行光合作用并为植物生长发育提供能量的细胞器。Wu等人[3]采用白鸡冠白化叶片进行叶绿体结构观察,发现叶绿体结构异常,叶绿体呈现空腔,基质类囊体和片层结构在白鸡冠白化叶片受损,并鲜见淀粉粒。而这一叶绿体发育异常的现象同样也发生在其他物种中,如水稻[94]、拟南芥[95]等白化突变体。在本研究中,相较于正常叶片,花白叶片的叶绿体结构出现异常,叶绿体结构出现空腔,存在着没有明显的基质类囊体结构的现象。利用石蜡切片进行研究花白叶片组织结构,结果表明叶片组织的疏松度和叶片厚度显著低于正常叶片,推测是花白叶片中的叶绿体发育异常而导致组织结构发育受阻。
叶绿素对于植物叶色和植物光合作用来说是一个重要色素。Feng 等[88]发现温敏型白化茶树安吉白茶和光敏型白化茶树的黄金芽、御金香中所含叶绿素的含量都低于福鼎大白(正常品种)。在本研究中,花白茶树也得到相似的结果表明了花白叶片叶绿素 a、叶绿素 b、叶绿素 a+b的含量均显著低于正常叶片。猜测花白茶树表型与色素有关。本研究中花白叶片的儿茶素总量低于正常叶片,而氨基酸总量显著增加。推测花白叶片相关代谢途径紊乱,造成相关生化成分含量变化。
5.2 花白茶树转录组和蛋白组学分析
捕光复合体(Lhc)是高等植物天线系统的主要组成部分,而存在于类囊体膜中的Lhcb在保护植物免受蛋白酶降解方面起着至关重要的作用。叶绿素含量与LHCb mRNA积累量呈正相关[96]。本研究发现花白叶片的Lhcb1和Lhcb3表达含量显著低于正常叶片(图4-6B)。叶绿体发育异常(图4-3B)与色素生物合成缺陷有关。这在水稻突变体lta1 和tcd5 得到了验证。本研究结果发现(图4-6A)花白叶片中编码原叶绿素氧化还原酶(POR)基因的下调。Griffiths等人发现POR下调导致叶绿素a的积累减少,叶绿素a是由严格依赖于光的POR酶介导的原叶绿素a的光还原产生的。原叶绿素a再通过进一步酯化和修饰的反应来产生叶绿素a和叶绿素b[49]。编码PorA的POR基因的异位过表达被证实可以恢复porB-1porc-1双零突变拟南芥的大量叶绿素积累,该突变体在叶绿体中具有未堆叠的类囊体膜,并表现出幼苗致死的黄色表型[97, 98]。POR在光敏白化茶品种白鸡冠中表达下调[3]。同样地,在温敏型的茶树小雪芽中,Li等[11]发现了两条POR基因均下调。因此,推测花白茶树的叶绿素含量降低与低表达的Lhcb1、Lhcb3和POR有关。
CHS是类黄酮途径中的第一个酶[99, 100]。矮牵牛和牵牛花中的CHI基因敲除突变减少了类黄酮的生产[101]。有研究表明茶树中编码类黄酮3’,5’-羟化酶的基因F3‘5’H是儿茶素(GC、EGC和EGCG)生物合成的重要调控因子[102]。ANR酶将花青素、花青素和飞雀素转化为EC和EGC,相应地将ANR基因进行过度表达会增加EC和EGC的含量[101]。检测花白茶树的差异基因的结果表明,白化叶片的CHS、CHI、F3‘5’H和ANR表达低于正常叶片。在检测花白茶树中的差异蛋白的结果也发现(表4-5),类黄酮的途径中的CHI酶在白化叶片含量是正常叶片的0.84倍,ANR酶的含量是正常叶片的0.81倍。白化叶片中CHI酶和ANR酶都下调,相应地,检测花白茶树的多酚含量结果显示白化叶片的儿茶素类含量大幅度降低,其中发现EC含量和EGC含量在白化叶片分别下降了30%和46%(表4-1)。花白叶片中儿茶素的降低,推测是CHS、CHI、F3‘5’H和ANR下调有关,以及ANR酶和CHI酶含量降低。此外,MYB和bHLH转录因子能在植物的次级代谢中发挥着重要的作用,尤其是在类黄酮代谢途径中[103, 104]。转录因子的分析结果中发现MYB和bHLH的转录家族在白化叶片和正常叶片具有显著变化(图4-8),推测白化叶片中多酚含量降低与MYB和bHLH的转录家族变化有关。
检测花白叶片和正常对照的游离氨基酸结果发现花白叶片其含量增加,尤其是谷氨酸和茶氨酸的含量高于正常叶片 (表4-2)。茶氨酸合成酶(TS)基因在白化叶片中的表达没有明显增加,然而谷氨酰胺合成酶(GS)基因表达含量则高于正常对照的表达含量(图4-6A)。GS序列与TS序列高度同源[105],在各种各样的植物中,如果有合适的底物,它可以合成茶氨酸。推测谷氨酸和茶氨酸含量增加是由于谷氨酰胺合成酶(GS)基因表达显著上调。在蛋白组学中,检测到了参与氮代谢和硫代谢的差异蛋白3 -磷酸甘油酸脱氢酶。有研究发现3 -磷酸甘油酸脱氢酶能够参与丝氨酸的磷酸化进而影响丝氨酸的生物合成途径,故表明其脱氢酶在丝氨酸合成中发挥着重要的作用[106]。
当拟南芥处于逆境中,氧化磷酸化可提供相应的能量[107]。本研究发现花白茶树中的花白叶片中参与氧化磷酸化、内质网蛋白加工和核糖体生物合成的酶有所积累(表4-5),这可能是由于花白叶片为了保证正常生长而必须增强相应的与生长发育相关的代谢途径,使得叶片得到更多的能量。
5.3 花白茶树的Clp蛋白酶复合体
小麦中VRS1中一个磷酸位点发生突变,会改变小麦的性状[108]。有研究表明,磷酸化位点可调控基因转录功能[109]。在本研究中,发现花白叶片中的ClpP5比正常叶片多了两个丝氨酸,改变了磷酸位点。叶绿体 Clp 蛋白酶复合体中的ClpP5亚基蛋白酶基因突变导致基因功能缺失会导致玉米幼苗叶片黄化[45]。推测花白叶片ClpP5可能会改变其功能或结构,从而导致茶树呈现白化性状。
ClpP5是Clp蛋白酶复合体的一个亚基,Clp蛋白表达水平降低会对Clp蛋白酶复合体组成和叶绿体蛋白稳态的影响。烟草NtEtClpP6突变体,降低了50%的ClpP6丰度,导致其他P-环和R-环组件(ClpP3、ClpP5、ClpR1和ClpR3等)的蛋白质丰度降低。推测ClpP5蛋白发生变化,相应的整个Clp复合体也会收到影响,改变了叶绿体蛋白的稳态[110]。
图5-1是参考拟南芥文献中所推测出的茶树中Clp蛋白酶复合体相关的机制。Clp蛋白酶复合体的主要功能是降解植物质体中受损和植物所不需要的蛋白[111, 112],若当复合体中的蛋白受到抑制或者发生突变的情况下,会影响着植物表型的变化,会使得植株叶色变为淡绿色甚至是全白,植株生长速度呈现出缓慢且矮小[113-115]。当ClpC蛋白受到影响(基因突变,表达抑制)也会影响着光合色素相关的蛋白,影响着类胡萝卜和叶绿素合成途径中的蛋白,其中包含着PSY(八氢番茄红素合成酶)[116]和CAO(脱植基叶绿素a氧化酶)[117],两者的蛋白含量会增加,影响着后续的色素的含量合成。相应地,当Clp蛋白酶复合体受到影响时,会破坏细胞内的蛋白稳态进而会影响着光合作用相关的蛋白和RNA修饰的蛋白[118]。推测Clp蛋白酶复合体与叶绿体发育、植株的生长发育有着很大的关联。

图5-1 茶树Clp复合体相关机制
Fig.5-1 Presumed mechanism of Clp complex in tea plant
花白茶树成为观赏植物或开发茶叶新产品存在着一定的市场,同样地也可成为研究叶绿体发育相关、核质基因突变等的理想材料。本论文分析花白茶树中花白叶片和正常对照的生理生化研究,并阐述花白茶树在分子层面上的变化来进行探究其相关的分子机理,为培育新品种的茶树以及与茶树相关的分子育种奠定了理论性的基础。
6.结论与展望
6.1 结论
本研究以黄山市耿城镇辅村茶园发现的花白茶树单株为研究材料,分析花白茶树花白叶片和正常对照的生理生化差异,利用转录组(RNA-seq)和蛋白组分析技术获得花白茶树中花白叶片与正常对照的差异基因和差异蛋白,并对影响叶绿体发育的Clp蛋白酶复合体在花白茶树中的表达进行分析,并利用PCR克隆Clp蛋白酶复合体在花白茶树中的差异表达基因。本研究取得的主要结果如下:
从叶片组织结构分析发现花白叶片的叶绿体结构发育异常,且呈现空腔。同时,花白叶片的组织结构疏松。相较于正常叶片,花白叶片叶绿素、儿茶素类含量减少,而氨基酸含量积累。在茶氨酸生物合成途径中,花白叶片的相关基因表达高于正常叶片。而在儿茶素和叶绿素生物合成途径中基因表达趋势则相反。相较于正常对照,花白叶片中儿茶素途径中相关蛋白含量显著降低。而维持生长发育所需的能量相关蛋白却升高。通过从花白叶片和正常叶片克隆出基因的序列分析后,发现CsClpP5基因在花白叶片中存在两个丝氨酸的位点突变,由分析发现该突变改变了该蛋白的磷酸化位点。从而猜测CsClpP5的突变,可能与茶树产生花白表型有关。
6.2 展望
花白茶树作为观赏性植株或作为茶叶新产品具有巨大的潜力,研究其白化机理在一定程度上具有重要意义。后续将CsClpP5 基因进行功能验证,研究CsPOR (NADPH-原叶绿素氧化还原酶)基因等差异基因。探索相关差异蛋白以及差异基因对其花白茶树中白化症状的影响,从而更深层次地进行研究其白化的分子机理。

1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:1158,如若转载,请注明出处:https://www.447766.cn/chachong/135652.html,