苹婆木属于梧桐科,苹婆树冠浓密、宽阔,且叶常绿不易落叶,是一种遮荫性能很好的行道树,而且树形美观,苹婆花花序大白里透红,每年可开花结实两次,蓇葖果放射状排列,成熟时呈鲜红色,因而具有很好的观赏性;而苹婆是原产于我国的乡土树种,并在广西有分布,具有生态适应性强,是生态、经济效益兼备的经济树林[1]。苹婆无论生长在石山或土山的谷地甚至在山坡上的石穴上都能生长良好,其果、材、皮具有较高的经济利用价值,在广西石灰岩山区有“木本粮食之称”[2]。可惜结实率不高;如能注意选育种、并且提高结实率,是一种值得提倡具有很高经济价值的木本粮食植物。苹婆全身是宝,特别是苹婆的叶子,是白头叶猴的主要食物,但是前人对苹婆的研究仅仅是在生长特性、繁殖方式、栽培管理等方面有初步的探讨。所以本文旨在对研究崇左地区苹婆的光合特性,不仅可以了解它的生理生态特性,而且对掌握苹婆的光合生产力等问题具有重要的意义,更为苹婆的栽培生产提供了理论参考,使其在园林植物配植中能得到更好的应用[3]。
1材料与方法
1.1实验材料
以常见景观植物苹婆作为研究材料,采用Yaxin-1102便携式光合蒸腾仪对苹婆的光合作用日变化特性进行测定。
1.2光合作用测定方法
选定生长势相近且无病虫害的植株作为供试材料。在晴朗、无云、强日光的天气条件(12月份、1月份、2月份,每个月重复3天做实验。)从8:00至18:00每隔l h测定一次光合速率日变化,测定时选取植株苹婆充分见光的完整成熟叶片作为测定材料。每次测定每片样叶重复计数5次[4],每小时完成25个气体交换光合值。测得9天的光合参数数据后,取均值进行统计分析[5-6]。研究指标包括光合有效辐射(PAR、μmol/m2.s)、净光合速率值(P n、molCO2/m2·s)、气孔导度(G s、mmolH20/m2.s)、胞间C02浓度(C i、μmol/mol)、蒸腾速率(Tr、mmolH20/m2.s)以及空气相对湿度(RH、%)、空气温度(Ta、0C)、空气C02浓度(Ca、μmol/mol)等。
1.3数据处理与分析
叶片瞬时水分利用效率(WUE,μmol/mmol)、光能利用率(LUE,mol/mmol)和气孔限制(Ls)分别用公式计算,即:(1)WUE=Pn/Tr;(2)LUE=Pn/PAR;(3)Ls=1-Ci/Ca,Ca是大气中的CO2摩尔分数。光合作用数据处理方法:测得9天的光合参数数据(去掉差异很大的数据组),做各个指标图,数据处理采用Excel软件完成。
2结果与分析
对苹婆冬季光合作用日变化特征研究先逐一分析各种环境因子,其次是分析内部因子,最后归纳影响苹婆光合特性的结论。
2.1苹婆环境因子日变化特征分析
如图1表示表明PAR呈现先升高后下降的单峰型曲线,光照强度较高出现在10:00-15:00时段,其平均值为1173.032μmol/m2.s;在11:00-12:00时段PAR逐渐升高,并且在14:00时出现一个微单峰趋势,峰值是1513.528μmol/m2.s;随后随时间变化呈下降趋势,且一天内下降趋势不同。
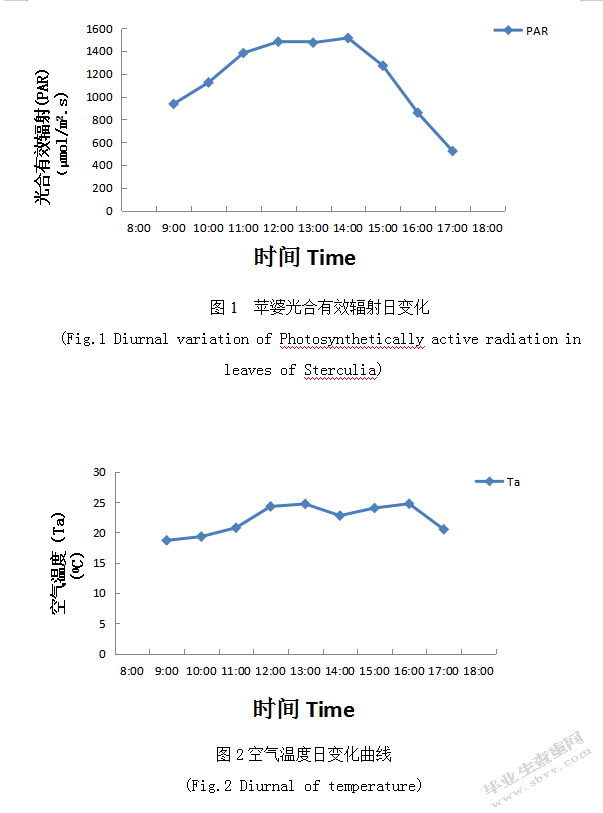

如图2表示Ta是出现微双峰型,在12:00和16:00各出现一个峰值,并且峰值基本相同;在14:00出现一个低谷,值是22.77997μmol/m2.s;14:00前表明Ta伴随着PAR升高而出现缓慢升高趋势,但是超过14:00后PAR逐渐减弱,而Ta缓慢升高直至16:00出现一个微峰值;16:00后与PAR日变化基本相同。
如图3表示RH日变化呈倒钟型,并且与Ta和PAR日变化呈负相关的关系。RH在早上出现最大值,其值为44.8%,在9:00-13:00时段之所以出现下降较快趋势,其主要原因是Ta缓慢升高导致空气密度下降,空气密度下降是Rhin降低一个重要原因,除此以外白天植物进行光合作用吸收CO2也是Ca降低一个重要原因[7]。
如图4表示CO2in日变化与RH日变化基本一致,在早上9:00出现最大值,值为464.16670C,之后随时间变化呈基本不变趋势,一天内变化的幅度基本相同。
因此影响光合特性的环境因素中,PAR是主要的驱动因子,PAR变化引起了Ta发生变化并且两者呈正相关型,但与RH和CO2in呈负相关型。植物光合作用日变化受各种环境因子影响,尤其是光照强度和温度的影响[8],诱发植物叶片光合作用的光抑制是因为植物叶片吸收的光能超过其转换能力时。PAR和Ca日变化如上图示:PAR出现午间高峰型单峰曲线趋势,超过14:00后呈下降较快趋势;Ca在9:00时出现最大值后呈下降趋势,并且在10:00-13:00时段缓慢上升趋势,13:00后呈缓慢下降趋势,故猜测Ca可能是影响苹婆的净光合速率主要环境因子。
2.2苹婆净光合速率日变化
植物光合作用是植物生产过程中物质积累与生理代谢的基本过程,也是分析环境因素影响植物生长和代谢的重要手段。一般条件下,栽培植物光合作用日变化均有规律可循,变化曲线呈双峰型或单峰型,单峰型中午光合速率最高,双峰型上、下午各有一高峰[9]。在自然条件下,植物的光合作用在中午降低是普遍现象[10],即光合午休,它是植物在长期进化过程中适应干旱环境而产生的一种生理现象,对植物节水与生存有利[11]。在自然环境中,植物光合作用一般在中午普遍降低的现象[12],它是适应严峻天气产生一种生理现象。如图5表明Pn日变化呈现先增加后下降的单峰型曲线,且在12:00达到峰值,即1.6336mmol/m2.s;在下午13:00-15:00时段呈下降较快趋势,超过15:00之后Pn日变化值与早上8:00-9:00时段Pn日变化值基本相同。该曲线说明了植物在午间积累的有机物最多,早晚有机物积累数量相对少。
2.3苹婆气孔导度日变化
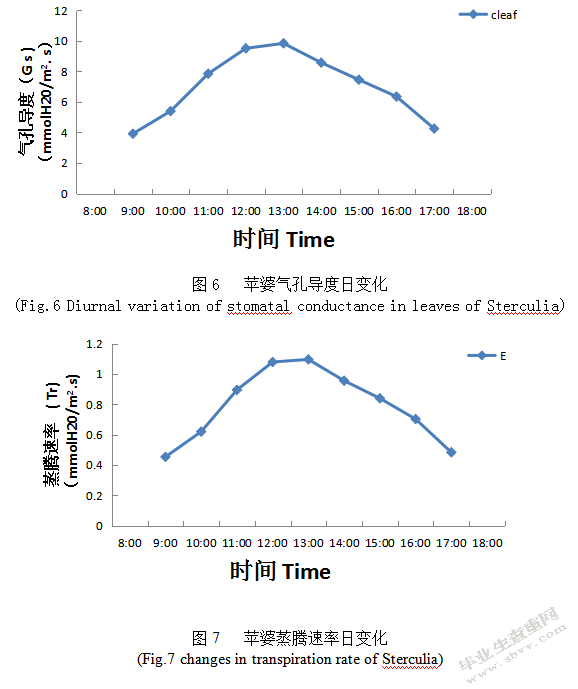
气孔是植物与外界环境进行水汽和C02交换的通道,植物叶片的光合作用和蒸腾作用随气孔的变化而变化,而气孔闭合被胞间C02浓度、空气相对湿度及温度等因素的影响[13]。由图6可知,Gs日变化和Pn日变化曲线基本一致,呈先增加后下降的单峰曲线,在10:00-13:00时段Gs长时间保持较高水平现象,且在13:00出现一个高峰,峰值为9.8281 mmol/m2.s;超过13:00后Gs日变化呈下降幅度较快趋势,且在17:00时Gs日变化趋势与早上基本相同。午间气孔导度之所以出现升高现象,其原因可能是叶温对Gs影响非常大,在合适的光强下能促进气孔开张,则气孔阻力降低,Gs增大;在土壤水分充足时,植物为避免高温灼伤叶片,将通过蒸腾作用带走叶片热量,增大气孔开度[14],有利于植物适应炎热的环境。
2.4苹婆蒸腾速率日变化
蒸腾是植物水分代谢重要的生理过程,植物通过蒸腾作用促进根部吸收矿物质、促进矿物质运输,还可以避免高温灼伤叶片,通过蒸腾作用带走叶面的热量,调节叶面温度、供应光合作用所需要的水分、调节气孔开度等,与植物净光合速率关系密切。由图7可知,Tr日变化呈先上升后下降的单峰型曲线,在12:00出现一个高峰,峰值为1.07838mmol/m2.s,且苹婆Tr与Gs日变化基本相同;如果苹婆气孔导度开放越大、则Tr日变化就越高。中午植物散失水分最快主要原因是12:00-13:00是光合有效辐射最强,大气温度最高,此时叶面温度升高,为了防止高温灼伤叶片,植物就要加强进行蒸腾作用带走叶面的热量,调节叶面温度。
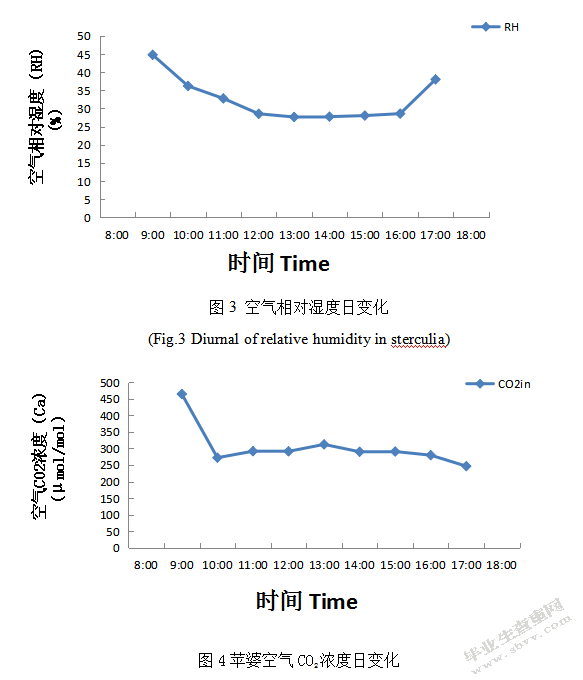
2.5胞间C02浓度日变化
植物气孔导度越大,进入细胞的二氧化碳就越多,二氧化碳是光合作用的原料,所以胞间二氧化碳就会越来越少,证明参与光合作用反应掉的二氧化碳越多,净光合速率就越大。由图8可知,苹婆冬季Ci日变化差别较大。苹婆冬季Ci与Pn日变化关系基本相反,Ci早晚低,中间呈上升趋势,但在10:00-16:00时段出现较快上升现象,且16:00时出现最大值204.8392μmol/mol;在16:00后呈迅速下降趋势,且与11:00变化基本相持平趋势。
2.6气孔限制值日变化
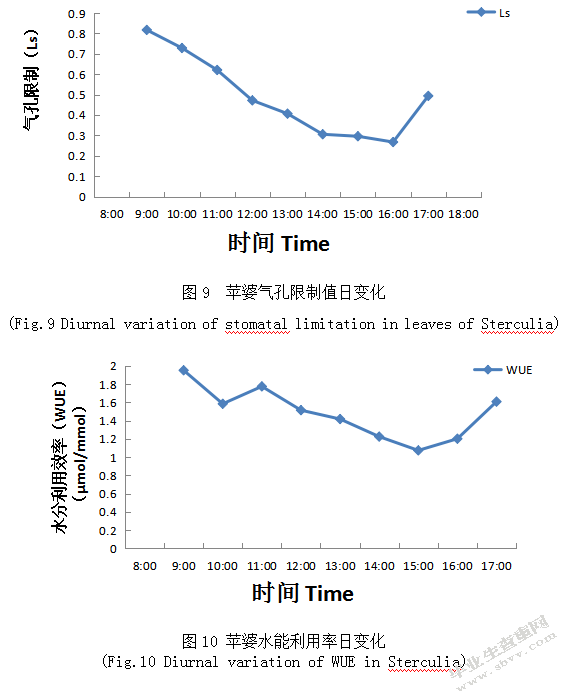
引起Pn减弱的原因有两个,其一是气孔限制,指气孔导度降低,进入气孔C02减少,不能满足光合作用的基本要求,其二非气孔限制,指叶片温度的增高,叶绿体各种酶活性降低,导致叶片光合作用能力降低。如图9表明9:00-18:00 Ls发生显著的差异,在10:00-12:00时段光合有效辐射逐渐增强,且叶面气温、蒸腾速率日变化值、净光速率和胞间C02浓度增强、气孔限制值下降;在13:00-14:00时段胞间C02浓度呈升高趋势,但是气孔导度出现下降较快幅度,所以气孔限制值下降;在14:00-16:00时段胞间C02浓度趋势基本不变、气孔限制值也保持不变;超过16:00之后随着光合有效辐射减弱,净光合速率也下降,则胞间C02浓度下降伴随着气孔限制值升高。
2.7水能利用率日变化
绿色植物利用光能将水和二氧化碳等无机物转化成有机物储存在植物体内并释放氧气,水是植物进行光合作用的原料之一,植物水能利用率WUE表明了植物经济用水能力。在相同的外在条件下,如果Pn值和WUE值相对较大,说明植物的水分利用效率高,反之则说明其水分利用效率低[15]。如图10表示苹婆WUE日变化是呈倒钟型,在10:00出现一个低谷,且在11点出现一个微峰,之后一直出现缓慢下降趋势,直至15:00时出现升高趋势。表明在10:00-11:00时段PAR升高,RH较湿润,此时Ta较低,因此WUE升高,但是随着PAR和RH升高,WUE日逐渐下降。在15:00-17:00时段,PAR显著下降同时Ta基本不变,此时RH开始上升,引起WUE逐渐升高
2.8光能能利用率日变化
绿色植物利用光能将水和二氧化碳等无机物转化成有机物储存在植物体内并释放氧气,光能能利用率表征植物能源物质来源。在相同的外在条件下,如果Pn值和LUE值相对较大,说明植物的光能分利用效率高,反之则说明其光能利用效率低。如图11表示植物在10:00-11:00时光能利用效率达到峰值,值为0.00115mol/mol。在11:00-13:00时段LUE出现缓慢下降趋势,可能引起的原因是PAR缓慢增强;在13:00-15:00时段LUE值下降较快,是因为光合有效辐射增强,叶面气温大,蒸腾速率日变化值也较强,最终导致LUE值下降较快;15:00后LUE值显著上升。
3结论
自然条件下,植物光合作用的日变化曲线存在两种类型:即单峰型,午间光合速率最高;双峰型,上、下午各有一个高峰。由于苹婆的光合速率日变化为单峰型,在12:00达到峰值,即1.6336mmol/m2.s。在本试验中,苹婆在自然条件下净光合速率与蒸腾速率、气孔导度呈正相关,与胞间C02浓度呈负相关,光合速率和气孔导度的峰形相近,气孔导度与光合速率呈正相关,苹婆光合作用日变化是生态环境因子综合作用的结果。
对苹婆光合特性探索尚需今后深入研究,希望借此抛砖引玉,引起更多人的关注,以及对园林设计中科学合理的植物配植的进一步研究,为苹婆的栽培生产提供了理论参考,使其在园林植物配植中能得到更好的应用。
4文献参考:
[1]冯文星,徐雪荣,雷新涛.极具开发潜力的热带干果果树——苹婆[J].中国热带农业,2006(06):19-20.
[2]朱宏光,农友,苏建苗,王家妍,李燕群,刘正富,赵金龙,林建勇,温远光.广西热带岩溶区苹婆幼树光合蒸腾特性研究[J].安徽农业科学,2011,39(13):7751-7752+7806.
[3]尤扬,陈太勇,贾文庆,宁晓明.柳叶榕叶片秋季光合日变化初探[J].贵州农业科学,2009,37(10):170-171+174.
[4]靳甜甜,傅伯杰,刘建国,胡婵娟,苏常红,刘宇.不同坡位沙棘光合日变化及其主要环境因子[N].生态学报,2013,31(7):1783-1793.
[5]李怡,刘发民,宋耀选,李毅.柽柳叶片光合速率日变化特征的研究[J].安徽农业科学,2008(18):7559-7560+7563.
[6]金则新,柯世省.云锦杜鹃叶片光合作用日变化特征[J].植物研究,2004,(4).
[7]张明农,江力.水稻光合特性的初探.安徽大学学报(自然科学版),1996,20(1):88~94.
[8]张明农,江力.水稻光合特性的初探.安徽大学学报(自然科学版),1996,20(1):88~94.
[9]邹琦.作物抗旱生理牛态研究.济南:山东科学技术出版社,1994.155—242.
[10]许大全,李德耀,沈允刚,等.田间小麦叶片光合作用“午睡”现象研究[J].植物生理学报,1984(10):269—276.
[11]张建新.气孔对水分利用的调节——Cowan和Farqu—har的气孔调节最优化理论[J].植物生理学通讯,1986(4):12-17.
[12]巨霞.不同类型油菜品种叶片的光合日变化特性[J].贵州农业科学,2013,41(07):31-35.
[13]邵玉文,韩梅,韩忠明,孔伟伟,杨利民.不同生境条件下黄岑光合日变化与环境因子的关系[J].生态学报,2008(3):1470—1477.
[14]刘玉玲,刘信凯,谌光晖,黄万坚.杜鹃红山茶与3个山茶栽培品种光合日变化特性研究[J].广东林业科技,2011,27(01):17-22.
[15]靳甜甜,傅伯杰,刘国华,胡婵娟,苏常红,刘宇.不同坡位沙棘光合日变化及其主要环境因子[J].生态学报,2011,31(07):1783-1793.
下载提示:
1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:写文章小能手,如若转载,请注明出处:https://www.447766.cn/chachong/12630.html,