1引言
辣椒(Capsicum frutescens L.)为茄科植物,也被称之为番椒、牛角椒、长辣椒、菜椒等,未成熟时颜色为绿色,成熟后颜色变为赤红色、黄色或绀青,以红色最为常见,常见的形状为圆锥形或长圆形[1]。它不仅是重要的蔬菜和调味品,具有极高的营养价值,补充人体需要的碳水化合物,维生素及钙、磷、铁矿物质等,同时还可入药,具有很好的保健功效,如驱寒止痛等功效[2]。然而,在辣椒广泛推广种植的同时,大面积种植辣椒也面临着各种病虫害以及疫病的威胁,造成辣椒的减产甚至绝收。使用化学药剂不能完全解决问题,反而会给环境和人类健康带来不可逆的危害。因此,防止辣椒疫病以及病虫害的关键是改良筛选抗病品种,提高辣椒自身的抗病性[3]。研究辣椒的天然免疫系统,是改良辣椒品种关键的知识。
1.1植物的抗病机理
1.1.1植物的天然免疫系统
植物生长和繁殖过程不仅面临着严峻的生态环境的挑战,同时还会受到各种病虫害以及疫病的侵染,但在长期的生物进化进程中,植物逐渐产生众多复杂合理的保护机制来抵御外界病原的入侵,上述机制如同植物免疫系统。上述系统通常被划分成不同层次,首先第一个层次是由病原体有关分子模式所催生的免疫反应(PTI),植物模式分辨受体利用辨别和病原体相关的分子形式,刺激活化促丝裂原活化蛋白激酶信号通路,进而在植物内出现初期的应答反应,应用范围广泛,效果显著,可分辨与响应大部分类微生物,比如细菌鞭毛蛋白、脂多糖等非致病菌的大部分微生物,进而立即触发活性氧爆发、超敏反应、植物内抗毒素的形成和部分有关抗病基因的表达上述基础免疫。此类防御反应可以高效的限制病原菌的生长和控制疾病的进一步发展,这与动物的自然免疫相似。第二个层次是通过效应因子触发型免疫(ETI)来重塑植物的抗性,病原菌形成效应因子,促使众多因子蛋白被传送到植物细胞质内和宿主蛋白彼此影响,限制基础免疫反应,进而在植物内出现众多病原体,因此就和哺乳动物的适应性免疫类似。此外使用PTI与ETI,把双方融合起来,可以高效的增加植物抗菌谱,优化植物ETI抗性[4-5]。
1.1.2植物的抗病反应特征
细胞的衰老死亡贯穿于植物整个生长发育过程之中,随着不断的深入研究,细胞死亡被划分成两类:坏死和细胞程序性死亡(programmed cell death,PHD)。二十世纪末期,Greenberg[6]等最先表明植物细胞程序性死亡的精准定义,该专家分析得知,即便不存在病原体侵染,拟南芥acd2突变体植株依然能自主产生病斑,且具备动物PCD的突出特点。细胞程序性死亡通常出现在生物发育时期,其需要信号或众多分子参加,是由基因影响的细胞自主的有序的死亡形式[7]。在细胞出现此类死亡时,细胞质压缩,细胞表层皱缩,染色体有规律的断裂成大小不均匀的碎片,在开展琼脂糖凝胶电泳时可查看到DNA“梯状”条带,此外肉眼查看到众多病斑[8]。PCD不仅在植物生长发育过程中有重要作用,而且在抵御外界胁迫方面以及与植物的超敏反应都有着非常密切的关系[9]。超敏反应(hypersensitive response,HR)概念最先由Stakman指出,是植物因为病原体入侵而出现自主局部细胞死亡,上述反应一般出现明显的PCD特征[10]。
1.2硫氧还蛋白
硫氧还蛋白(thioredoxin,Trx)普遍出现在动物、植物、细菌、酵母中,是一种分子量约为12kD的酸性小蛋白。
1.2.1结构特征
在硫氧还蛋白中存在很多的氨基酸残基,随着研究的深入和生物技术的发展,研究证明这些氨基酸残基对其蛋白结构和催化活性的重要性是不可忽视的。我们已经可以从植物中分离得到Trx,植物内大多数Trx都包含相对保守的外露活性中心,包含两个半胱氨酸残基。氧化态Trx的活性中心出现二硫键(S-S),包含两个胱氨酸残基,可以被NADPH与NADP/Trx还原酶体系、二硫苏糖醇(DTT)或还原态淀粉酶抑制蛋白(ASI-red)所还原。活性中心一边存在比较平缓的疏水区,此区是Trx和靶蛋白的融合位点,其特异性结合取决于活性中心周围的氨基酸序列的变化。经核磁共振等生物技术手段分析,全部硫氧还蛋白的二级结构均具有五个保守β链,被四个α-螺旋围绕,也就是β1,α1,β2,α2,β3,α3,β4,β5与α4,一般被叫做硫氧还蛋白折叠。在第二个β链末端以及α-螺旋的开端是硫氧还蛋白的活性位点。伴随硫氧还蛋白结构的改变,两个硫分子彼此间距离变大,半胱氨酸N-端旋转,最后出现活性中心二硫键的还原。由于N-端转运肽的体积不同,这种结构上的差异使得硫氧还蛋白被分为Trxm、Trxf与Trxh,其中Trxf和Trxm的差异在于在N-端处的不同,Trxf和Trxm相比更长,且在C-端出现保守的半胱氨酸,此结构和动物硫氧还蛋白类似[11-13]。
1.2.2生物学功能
由于Trx中双硫键(S-S)的存在,其氧化还原反应可高效传递电子,这种氧化还原反应具有可恢复和快速的特点,作用范围广泛,所以Trx可利用管控S-S氧化态结构或S-H基团的形式来调整管控蛋白质的作用。基于当前的分析进度可知,Trx已发现存在于十几种生化反应体系中,在植物体内的作用举足轻重[14]。植物内Trx被寻找时出现在菠菜叶绿体内,被当做和光合作用有关,然而伴随分析的加深,不只在叶绿体内,在细胞质与线粒体内也出现硫氧还蛋白,其在结构以及功能上出现细微的不同[15]。动物体内TRX种类明显更少,但其作用仍然不可忽视。
在夏德习[16]的实验研究中,选择了几种典型的逆境,研究分析了硫氧还蛋白基因TRXm1的表达特性,结果表明TRXm1基因在NaCl、NaHCO3、ABA、H2 O2处理下都有一定的应答反应,而对NaHCO3、ABA、H2 O2逆境的胁迫的应答关系又大于NaCl,然后通过转基因技术将硫氧还蛋白基因TRXm1导入拟南芥中使其过量表达,比较野生型与转基因拟南芥的生长情况,后者生长较快,因此我们就可以知道硫氧还蛋白基因AtTRXm1的功能和环境逆境紧密关联。在种子萌发初期,h型硫氧还蛋白可以让水解酶抑制剂失活,此外影响激活α-淀粉酶、支链淀粉酶等的功能,出现在植物细胞质、内质网、线粒体内,此问题表示Trxh的功能是可当做种子萌发的信息。在萌发时期,不同氧还原物质变化明显,Trxh在此时期功能更加显著,可以降解植株体内蛋白质等[17]。
1.3研究目的和意义
近年来,对于硫氧还蛋白的研究,不只模式植物拟南芥中得到了良好的发展,此外还从其余植物内分割出硫氧还蛋白基因,效果显著,对其内部结构,表达与功能进行详细全面的分析。对于硫氧化蛋白的结构与生物学功能等进行分析,为大众深入研究植物的抗逆性提供基础。本实验在克隆了一个辣椒硫氧化蛋白基因Trh9基础上,通过在辣椒叶片的瞬间表达,发现它能够引起过敏性细胞死亡,暗示该基因可能参与了辣椒应答病原菌的反应过程,为后续分析此基因在抗病反应内的作用提供帮助。
2材料与方法
2.1实验材料
本次实验研究采用质粒载体包含入门载体pDONR-207,目的载体3687,大肠杆菌菌株使用DH-10 B,农杆菌菌株是EHA105,实验所用辣椒品种是由福建农林大学辣椒育种团队提供并保存的Fj8。其它药品与试剂均由实验室提供。
2.2试验方法与步骤
2.2.1利用gateway技术构建Trh9基因及瞬间表达载体
首先以辣椒cDNA为模板,通过PCR技术分离并克隆Trh9基因,并进行测序。然后用Gateway中的BP反应方法,将全长的Trh9 cDNA构建到入门载体PDONR207上,提取Trh9-PDONR207的质粒,再用Gateway中LR反应的方法将Trh9-PDONR207构建到目的载体3687的植物表达载体中,即获得过表达载体35S:Cath9,然后用获得的过表达载体35S:Cath9转化农杆菌进行下一步实验。
2.2.1.1BP反应
1)pDONR-207是attP供体载体,通常使用在BP反应内创建入门载体,主要涵盖抗kan与抗cm筛选标记,此外包含ccdB基因,由于上述基因编码影响大肠杆菌DNA促旋酶的蛋白合成,因此限制标准大肠杆菌宿主的成熟。
2)BP反应体系

3)转化大肠杆菌实验
选取离心管,添加100μL感受态细胞,之后把BP反应的产物均转移到此管内,马上冰浴三十分钟。进而取出离心管,放置到42℃的恒温水浴锅中实施热激,时间九十秒。结束热激环节之后,马上添加到冰盒中,冰浴离心管内的细胞五分钟。结束之后添加800μLLB液体培养基(不含抗生素),以200r/min转速在37℃摇床内振摇1 h。之后制备包含卡那霉素的平板,吸取细菌悬浮液涂布于上,此时要格外关注不集结,吹干之后封闭,在37℃恒温箱内倒置培育基平板,时间是16-18 h。等到其中产生菌落,选择菌落放到液体培养基(含有卡那霉素)的离心管中,以200r/min转速在37℃摇床中培育三小时。
3)菌液PCR验证
等到大肠杆菌转化之后,开展PCR验证,引物是内侧特异性引物(划分成两组,一组中包含SRDX短肽,其他不包含)。
PCR反应程序是95℃预变性五分钟;94℃变性三十秒,此外完成三十五个循环;54℃退火5五十秒;72℃增加两分钟;最后72℃增加十分钟。对其开展PCR之后,把产物添加到1%琼脂糖凝胶电泳中,实施跑胶二十分钟,电压120 V,最终在凝胶成像仪上测试PCR产物且拍摄记录。
4)大肠杆菌质粒的提取
选取离心管,增加5mL正常菌液,添加到12000rpm离心机内进行离心,时间一分钟。离心结束之后清除上清液,之后在离心管内增加250μL含RNaseA的溶液P1,此时尽量摇晃,悬浮划分细菌内的细胞沉淀。此后增加P2溶液250μL,且缓慢摇晃,保证DNA片段不存在断裂,菌体全部裂解。在其中添加P3溶液350μL,且立即缓慢振荡,全面混合溶液,一直到其中存在白色沉淀物质。把其放到12000 rpm的离心机内等待十分钟,取出离心管,把上清液吸到吸附柱中。之后第二次放到12000 rpm下实施离心一分钟,舍弃收集管中的废液,留存收集管,将吸附柱放回远处。在其中增加漂洗液PW(包含无水乙醇)500μL实施洗涤,之后反复上述环节,清洗两次。之后将吸附柱放到收集管上,实施离心,速度12000 rpm,时间两分钟。之后,直接静置十分钟,尽可能去除其中剩余的乙醇,之后选取全新的离心管,放入吸附柱,此后在吸附膜的中心部分增加洗脱液EB大概40μL。最终,将离心管放置两分钟,此后在12000 rpm下离心两分钟,此刻包含质粒的溶液随之被筹集到离心管内。(为了提高质粒的回收效率,反复环节8。)
2.2.1.2LR反应
1)LR反应体系
细心混合液体,开始高速离心,之后在25℃温度内温浴过夜。
参考2.2.3.1-2方式开展大肠杆菌的转化,让得到具卡那霉素抗性的单克隆菌液。参考2.2.3.1-3方式,将单克隆菌液当做模板,使用内侧特异性引物开展PCR判定。PCR反应结束以后开展1%琼脂糖凝胶电泳跑胶20 min(120V),在成像仪上查找PCR产物此外拍照记录。依照测试结论选择阳性克隆开展后续环节。此后参考2.2.3.1-4方式提取大肠杆菌质粒,使用到后续的农杆菌转化测试中。
2.2.2农杆菌转化及PCR鉴定
2.2.2.1农杆菌转化实验
在具有100μL的农杆菌感受态细胞内增加质粒混合液(未含SRDX短肽)10μL,之后混合,冰浴三十分钟。把具有混合液的离心管放到液氮盒内冰冻,时间大概是三分钟,此后放到37℃水浴锅内,时间五分钟,开展热休克。选取离心管,增加500-1000μL未包含抗生素的液体LB培养基,缓慢缓和,之后放到28℃摇床,速度是200 r/min,振摇培育,时间是6 h。选取之前转化的菌液,主要在含有卡那霉素与利福平的LB平板上完成涂布,此后倒放在28℃恒温培养箱内2 d。等到产生菌落,挑选未连接的菌落至含有卡那霉素与利福平的液体LB培养基的离心管内,放到温度是28℃,速度是200 r/min的摇床中振摇过夜培育。反复以上环节进行含SRDX短肽的质粒混合液开展农杆菌的转化。
2.2.2.2菌液PCR验证
选择少数农杆菌菌液,在99℃温度下加热十五分钟,促使农杆菌的细胞壁破裂。之后使用内侧特异性引物鉴定PCR。PCR反应系统与程序同BP反应。均结束之后将扩增产物在1%琼脂糖凝胶内电泳20 min(120 V),在成像仪上查找PCR产物此外拍照记录。依照结论选择阳性克隆,扩增繁殖且储存,使用到后续的测试中。
2.2.3辣椒苗的处理及叶片的瞬间表达分析
2.2.3.1辣椒苗的处理
将Trh9-3687转化的农杆菌当做实验组,Trh9与PK7空载体是对照组,把上述两组的农杆菌放在培养基内过夜。在菌液吸光度是1.0时,把菌液放在8000 rpm的离心机中操作,舍弃上清,使用VIGS侵染液重悬沉淀,调整吸光度是0.8。之后使用重悬结束的农杆菌液对生长大概4周的辣椒苗叶片实施注射喷施。把辣椒苗放到室内培养且查看表型变动,等到后续测试。
2.2.3.2组织化学染色
DAB染色(二氨基联苯胺法):选取经过农杆菌与对照处理48 h的辣椒叶片10片,在清洁之后放到1 mg/mL且pH为3.8的DAB染色液中,在室温和无光照条件下处置8 h,此后使用95%的乙醇溶液在沸水浴内洗脱二十分钟,促使染液被全部洗掉,最终选择叶片放到显微镜下查看且拍摄。台盼蓝染色(Trypan blue):剪取通过农杆菌与对照处理的辣椒叶片10片,经过清洁之后放到台盼蓝染色液(乳酸:甘油:苯酚:水:1%台盼蓝:无水乙醇=1:1:1:1:0.1:4)中,在沸水中煮,时间五分钟。在室温和无光照环境中处理8 h,此后使用1%的水合氯醛在沸水浴中脱色,时间二十分钟,在蓝斑产生,脱色完成,选取叶片且进行详细观察,拍摄记录。
3结果与分析
3.1Trh9基因分离克隆的结果与分析
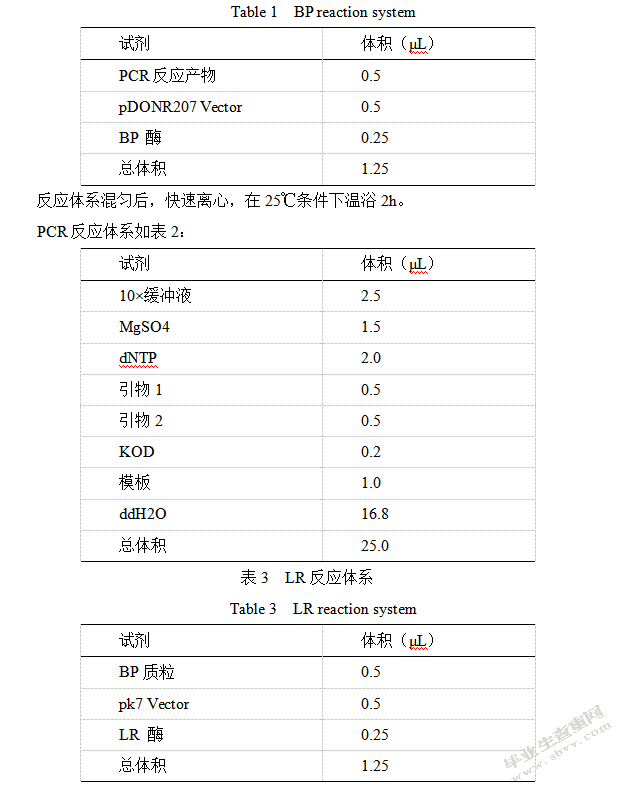
以辣椒的cDNA为模板,设计Trh9的特异扩增引物进行PCR扩增,得出长度大概是500 bp的PCR扩增片段(参考图1),把PCR产物以pDONR-207为载体转化大肠杆菌,选取质粒且开展测序。对最终结果开展序列比较,得知其和辣椒基因组数据库序列相同,也就可以得到我们需要的基因。
把基因序列添加到NCBI网站开展BLAST测试,最终结果参考图2,这说明该基因是一个硫氧还蛋白,编码氨基酸152个。我们对Trh9编码氨基酸的同源蛋白进行了比对分析(图3),结果发现辣椒(Capsicum annuum)Trh9与烟草(Nicotiana attenuata),番茄(Solanum lycopersicum),牵牛花(Ipomoea nil)同源性比较高,而且发现其保守的活性位点(WCGPC),说明它是Trx h的一个成员。
3.2构建Trh9基因的过表达载体
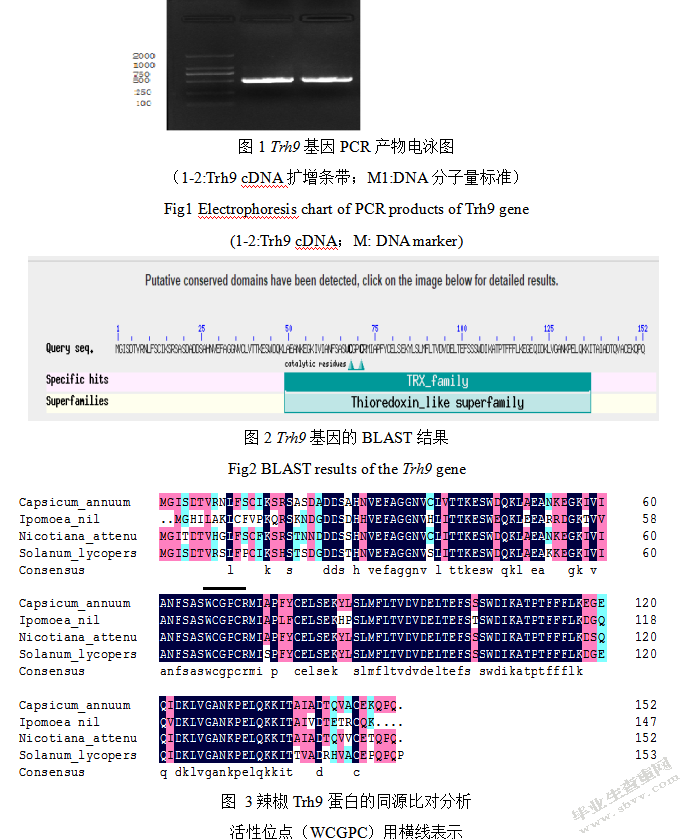
使用gateway技术对Trh9基因创建瞬间表达载体,利用BP与LR反应,把Trh9基因片段关联到pk 7载体。采用以上基因转化农杆菌,对转化产物实施PCR扩增,最终电泳结果图参考图4,根据内容可知PCR扩增产物的长度大概是500 bp,其和Trh9基因的长度符合,表示农杆菌转化顺利。
3.3构建Trh9-3687的过表达载体
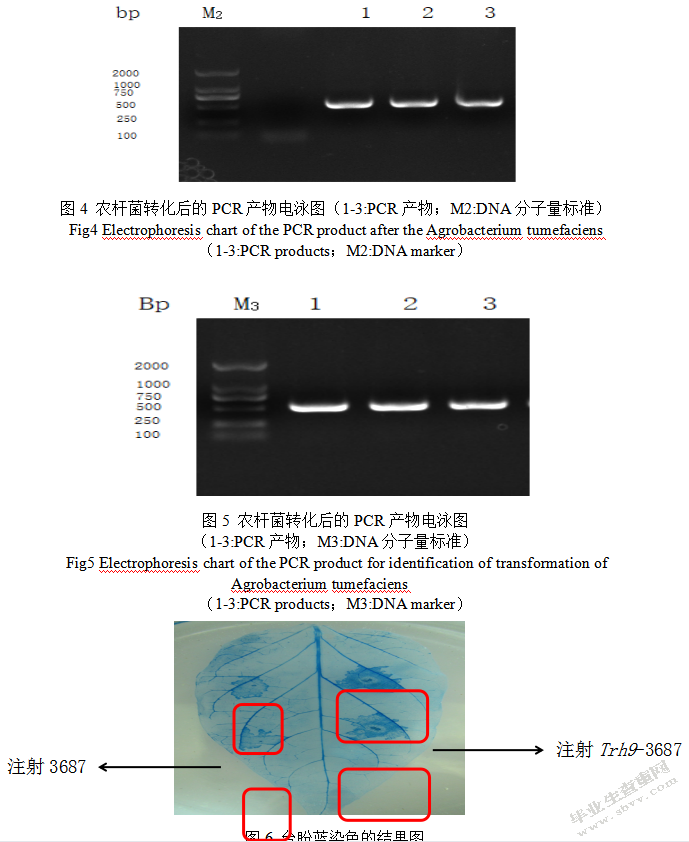
PCR扩增之后创建到3687载体内,转化结束之后,开展PCR验证,最终电泳结果图参考图5,根据内容可知PCR扩增产物符合预期,表示顺利得到Trh9-3687的瞬间表达载体。
3.4辣椒叶片组织化学染色结果与分析
植物细胞膜具备选择透过性,然而现已死亡的细胞就丧失此效能,通透性增提高,因此在使用台酚蓝染色时,一般植物细胞膜会阻碍染色液进入细胞染料,但是死亡的细胞并不会这样,从而细胞被染成蓝色,在图6中可以看出,注射Trh9-3687的辣椒叶片被大范围被染成蓝色,但是注射3687(对照)辣椒叶片只是在注射点被染色。
当植物遭受侵染之后,侵染点四周局部组织或细胞会马上坏死,进而避免病原菌扩散到其余没有被浸染的部分。此刻,植物细胞频繁死亡且释放出众多过氧化氢。DAB染色时,过氧化物酶处于细胞内,可以释放过氧化氢内的氧,此后二氨基联苯胺(DAB)全部氧化,产生金黄色沉淀。DAB染色情况参考图7,我们从图中可以看出注射Trh9-3687的辣椒叶片大范围被被染色,但是3687只在注射点周围被染色。
4讨论
4.1辣椒硫氧还蛋白的基因序列及功能保守性
用ExPasy网(http://www.expasy.org/)预测分析结果表明基因编码152个氨基酸。使用NCBI的保守功能区域研究程序来进行预估,结论指出该基因为I类Trx家族基因,此处编码蛋白具备硫氧还蛋白相对保守的活性中心序列胱氨酸-甘氨酸-脯氨酸-胱氨酸(CGPC)。在茄科植物以及非茄科植物的硫氧还蛋白基因Trh9基因序列的多重比较中,也发现其序列的保守性。硫氧还蛋白的作用贯穿植物生长过程始终,其中各个基因各司其职,基因之间的序列相似性很高,在不同生长发育阶段参与植物正常生理活动。
4.2 Trh9基因与辣椒的抗病反应相关
当前主要使用DAB与台盼蓝染色法对不同组别的辣椒叶片进行染色,依照最终结果可知,注射Trh9-3687的辣椒叶片经过染色后出现显著的过敏反应,染色突出,但是对照组染色效果不突出,表示Trh9基因表达遭受抑制会造成植物细胞程序性死亡,和其抗病性有相应的关系。
5结论
硫氧还蛋白基因家具备序列及功能的保守性,且与植物的抗病反应密切相关。我们利用Gateway技术构建Trh9基因及其过表达载体35S:Trh9,用其对辣椒叶片进行注射处理,结果表明Trh9可能会引发植物细胞过敏性程序死亡。
参考文献
[1]刘洋,龙应霞.辣椒研究现状及发展策略[J].黔南民族师范学院学报,2009,(3):39-43,54.
[2]高翔.辣椒的保健功能及其产品的开发研究[J].食品研究与开发,2004,(3):115-116.
[3]易图永,谢丙炎,张宝玺,等.辣椒疫病防治研究进展[J].中国蔬菜,2002,(5):52-55.
[4]陈英,谭碧玥,黄敏仁.植物天然免疫系统研究进展[J].南京林业大学学报(自然科学版),2012,(1):129-136.
[5]JONES JDG,DANGL JL.The plant immune system[J].7117(7117):323-329.
[6]GREENBERG J T,GUO A,KLESSING D F.Programmed cell death in plants:a pathogen-triggered response activated coordinately with multiple defense functions[J].1994,77551-563.
[7]BREUSEGEM F VAN,DAT J F.Reactive oxygen species in plant cell death.[J].2006,2(2):384-390.
[8]谭茂玉,沈法富,王峰翔.植物细胞程序性死亡研究进展[J].山东科学,2007,(2):17-23,28.
[9]孙鑫博,代小梅,王怡杰,等.植物细胞程序性死亡研究进展[J].生物技术通报,2010,(11):1-6.
[10]夏启中,等.与植物超敏反应(HR)相关的细胞编程性死亡[J].华中农业大学学报,2005,24(1):97—103.
[11]卫丽,黄晓书,李鹏坤,等.植物硫氧还蛋白研究进展[J].贵州农业科学,2006,(6):129-131,123.
[12]孙虎,薛保国,杨丽荣,等.植物硫氧还蛋白系统[J].基因组学与应用生物学,2010,(4):748-753.DOI:10.3969/gab.029.000748.
[13]Homlgren,A.Tioredoxin[J].Ann.Reu.Biochem.1985,(54):237-271.
[14]刘雷,尹钧.硫氧还蛋白的研究[J].东北农业大学学报,2003,(2):219-225.
[15]LAUGHNER B J,SEHNKE P C,FERL R J.A novel nuclear member of the thioredoxin superfamily[J].118987-996.
[16]夏德习,管清杰,金淑梅,等.拟南芥硫氧还蛋白M1型基因(AtTRX m1)与环境逆境之间的关系[J].分子植物育种,2007,(1):21-26.DOI:10.3969/j.issn.1672-416X.2007.01.004.
[17]Laloi C,Rayapuram N,Chartier Y,et al.Identification and characterization of a mitochondrial thioredoxin system in plants[J].Proceedings of the National Academy of Sciences of the United States of America,2001,(24):14144-14149.
下载提示:
1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:写文章小能手,如若转载,请注明出处:https://www.447766.cn/chachong/13917.html,