摘 要
鸡的卵泡选择与产蛋量高度相关,研究卵泡选择的分子机制可为提高其产蛋性能提供一定的理论依据。Wnt信号通路能够调控很多卵巢生长发育过程中的生理过程,如卵泡发育、排卵和黄体化。Wnt4是Wnt家族的成员,对雌性动物的生殖系统起到一定的调控作用。为了研究Wnt4对鸡卵泡选择的调控作用,本研究检测了Wnt4重组蛋白对鸡等级卵泡颗粒细胞中相关基因表达的影响作用,结果发现Wnt4重组蛋白能够显著上调FSHR和CTNNB1的表达,并对StAR和CYP11A1的表达有一定的促进作用。本研究的结果可为深入研究Wnt4的具体功能提供一定的研究基础,也可为进一步阐明鸡卵泡选择的分子机制提供参考。
关键词:鸡;卵泡选择;Wnt信号通路;Wnt4;qRT-PCR
1.文献综述
我国是最早开始驯养鸡的国家之一,在许多地方都发现有四千多年前人们所剩的鸡骨和所制的陶鸡。中国家鸡由古代的野鸡驯化而来,因品种优良,数量多,对世界产生了深远的影响。养禽业是饲养家禽的生产企业,在畜牧业中,养禽业作为我国最早的饲养活动之一,一直处于比较重要的地位[1]。它的优点也较为明显,较之饲养猪、牛、羊等家畜相比,饲养鸡生产周期短、饲料转化率高、投资少,这三个优点使得饲养鸡在我国非常普遍,但是我国养鸡业普遍生产规模较小,繁殖、饲养手段较为落后。随着社会经济的发展,人们的生活水平有了很大的提高,对食物的要求也逐渐提高。制约鸡产蛋量的关键因素包括卵泡选择和卵泡发育,因此研究卵泡的选择和发育机制显得尤为重要。本研究主要检测了Wnt4重组蛋白对鸡等级卵泡颗粒细胞相关基因表达的影响,可为阐明Wnt4的功能提供一定的研究基础,也可为提高鸡的产蛋性能和繁殖力提供一定的理论参考。
1.1鸡的繁殖特点
鸡作为禽类的一种,以卵生的方式繁衍后代,因此卵巢功能的正常执行是其卵泡生长发育和排卵等一系列生殖行为的基础。母鸡的生殖系统由卵巢和输卵管组成,但是一般只有左侧的卵巢和输卵管发育完全,并有生殖功能。鸡卵巢是一个不断发生规律性变化的器官,是雌性生殖系统中重要的结构器官,同时也是一种必需的内分泌器官,这对于卵巢排卵和卵泡增长具有重要意义[2]。鸡以特定节律(24h左右)排卵,而且在母鸡腹部内存在不同阶段的卵泡,卵生动物最重要的繁殖特点就是卵泡等级制度,这是包括他们远古祖先在内所有卵生动物的共同特征[3]。
1.1.1卵泡发育
鸡作为鸟纲动物的一种,它在胚胎发育的早期有一对对称的卵巢,但有,随着研究发现,很多鸟类在出生后右侧卵巢消失,只剩下的左侧卵巢能够发育成熟行使功能。母鸡的卵泡发育非常的复杂,需要受到其他器官的协调,且受到各种分泌因子的调控[4]。母鸡的卵泡按照功能形态的不同分为两大体系:等级前卵泡和等级卵泡。它的发育排卵遵循严格的等级制度。禽卵泡由卵母细胞组成,卵母细胞被一层颗粒细胞包围,颗粒细胞与卵母细胞质膜接触。卵泡膜细胞层包围颗粒细胞。在3种细胞类型中,旁分泌信号和相关的细胞内转录途径被激活,对于调节许多物种的卵泡发育至关重要。卵泡的增长是持续的,依次生长继续生长直至死亡或排卵。在开始生长时,卵泡数量取决于小卵泡池的大小,卵泡生长独立于促性腺激素,及卵巢内的因素,大卵泡的数量越来越多,依赖促性腺激素和外源性促性腺激素,在很大程度上降低了卵泡闭锁的发生率。卵泡发育是一个漫长而复杂的受管制的过程,包括定期的扩散和精确控制分化体细胞和生殖细胞。
1.1.2卵泡选择
高效产蛋母鸡的卵泡发育以组织良好的卵泡等级为特征。其他鸡的情况并非如此,如肉种鸡卵泡发育过度,生殖能力效率低下。卵母细胞的相互作用促进卵泡选择[5]。抗穆勒氏管激素(AMH)已被证明可以表示调节哺乳动物卵泡发育的速率。在母鸡上,卵泡群与哺乳动物在卵泡刺激素分泌的表达上相似,在前几年卵泡选择时间显著减少。在产蛋的高峰期,母鸡以特定节律从小黄卵泡中选择一个进入等级卵泡,迅速生长繁衍,这个过程就称为卵泡选择[5]。虽然在一个产蛋周期内只能有一个小黄卵泡进入等级卵泡,但是有6-12个直径为6-8mm的小黄卵泡存在于鸡的卵巢内部,这与卵泡的FSHR 的表达量有着密不可分的关系,FSHR的表达量越高,小黄卵泡越可能被选择,但是当这个小黄卵泡被选择后,它的FSHR的表达量会逐渐减少[6]。小白卵细胞会发育成小黄卵细胞,在这个过程中小黄卵细胞池中被不断地填补进被生长激活后的小白卵细胞。在鸡的卵巢中未被选择的小黄卵细胞并不会失去活性,这与其他的哺乳动物并不一样,未被选择的小黄卵细胞会供给下一个周期,重新经历一遍选择过程[7]。在一个发育完善的鸡卵巢中,只有几百个卵母细胞能发育成熟并被排出,这是在巨大的12000个基数情况下,剩下的卵母细胞就会失去发育能力,因此研究卵泡选择的分子机制对于充分开发禽类卵巢中的卵泡资源十分重要。
1.2 Wnt信号通路
1.2.1 Wnt信号通路简介
Wnt蛋白家族,由19种非可溶性分泌型糖蛋白组成,从组织学角度可分为经典型和非经典型[8]。经典型Wnt蛋白与低密度的跨膜蛋白结合,非经典Wnt蛋白不依赖β-连环蛋白的转录功能。Wnt信号通路可以调节控制机体疾病的发生和发育过程,是调控细胞增殖分化的关键环节[9]。Wnts在一般情况下,作为配体来参与信号转导,有15种或更多的受体或者协同受体来相互作用,它们的相互作用可以引发三种不同的信号转导[10]。
1.2.2 Wnt家族成员在卵泡发育中的作用
Wnt信号通路有助于卵巢和卵泡的发育。Wnt家信号通路能够参与多种生理过程,如胚胎的生长发育、排卵和代谢[11]。Wnt信号失调,从而导致基因表达过量,可能会引起基因突变,产生疾病[12]。Wnt信号通路可以阻碍脂肪细胞分化,此外Wnt信号通路还可以控制细胞的增值和凋亡。Wnt具有比较高的保守性,可以参与体内多种发育过程[13]。随着科学技术的发展,生物学家对Wnt家族在卵生动物遗传发育中的影响,已获得重大的突破,值得一提的是,Wnt家族和其表达的信号在其他的哺乳动物,例如人类、羊还有鼠类的黄体和发育卵泡中都有被检测到[14]。针对Wnt的功能研究发现,鸡卵泡的选择与发育与Wnt4的功能、转录调控有关[15]。
1.3 Wnt4
1.3.1Wnt4简介
Wnt4是Wnt家族成员的重要一员,它是是调节Wnt /β-catenin信号通路的关键蛋白,目前在小鼠、人、牛中都检测到它的存在[18]。Wnt4有研究证明其在人类和小鼠的早期胚胎发育中起着关键作用[16]。Wnt4可以调控胚胎卵巢的发育和分化,在哺乳动物的小卵泡中高度表达,此后逐渐减少,直到排卵[17]。在小鼠中,Wnt4参与了卵巢的发育生长和成熟,也可以调控雌性动物的性腺分化。Wnt4还能调控人的黄体化颗粒细胞中相关基因的表达。
1.3.2Wnt4在卵泡发育中的作用
Wnt4对雌性动物生殖系统的发育起关键作用[19]。目前,人们对Wnt4的注意力主要集中在对哺乳动物(尤其是人和鼠)上,尤其是Wnt4对卵巢发育的影响[20]。敲除Wnt4的小鼠,其卵母细胞的数量明显下降,生殖能力下降,生育能力受到阻碍[21]。鉴于Wnt信号通路的保守性,我们猜测, Wnt信号通路也能够参与禽类的卵泡发育,且Wnt4是调控卵泡发育所必不可少的[22]。
1.4本研究的目的及意义
Wnt4作为Wnt家族的成员,其功能的研究十分重要。本研究中,我们检测了Wnt4重组蛋白对鸡等级卵泡颗粒细胞中与卵泡选择和发育相关基因的表达影响,这可为进一步丰富对Wnt4功能的研究提供一定的实验数据,也可为进一步明确卵泡选择的分子机制提供一定的参考,在提高家禽经济效益上起到极其重要的作用[23]。
2.材料和方法
2.1实验材料
2.1.1实验动物
随机选取三只200日龄处于产蛋高峰期的母鸡(海兰褐壳蛋鸡)及时处死,取其所有卵泡按标准分好等级,用于培养卵泡颗粒细胞。
2.1.2实验器材
超低温冰箱、低温高速离心机、荧光定量PCR仪、PCR仪、稳压电泳仪、凝胶成像系统、超净工作台、电子天平、制冰机、高压蒸汽灭菌器、二氧化碳培养箱、微量高速离心机、移液器、电热恒温鼓风干燥箱、正置照相显微镜、细胞低速离心机、超纯水机、细胞培养板等实验耗材。
2.1.3实验试剂
Wnt4重组蛋白,M199培养基,胰酶,胎牛血清,青链霉素,Trizol,氯仿,异丙醇,75%乙醇, RNaseFreeH2O,DNAmarkers,SYBRPremixExTaqII 定量试剂盒,反转录试剂盒,II型胶原酶,胎牛血清,青链霉素,PBS等。
2.2实验方法
2.2.1鸡卵泡颗粒细胞的培养
(1)产蛋高峰期母鸡,使用颈静脉放血的方式杀死,用消毒的小剪刀完整取下卵巢,用消毒的大剪刀剖开腹部,投入盛有3%青链霉素PBS(实验前注意进行灭菌)的无菌大烧杯中,进入无菌间完成分离。注取卵巢时应注意保持卵泡的完整。
(2)选择直径12mm以上的各级卵泡,逐个取下放于盛有3%青链霉素PBS的灭菌平皿中,在超净工作台中,用左手拿尖头镊子,右手用尖头镊子将卵泡的外层膜挑开进行剥离,剥离后暴露基膜层,用尖头镊子剪破基膜层放出卵黄,为了免损伤颗粒层,在释放卵黄时动作的一定要轻柔,颗粒层中可能带有卵黄,为减少卵黄的含量可以洗涤几次,释放出卵黄后,将基膜层和颗粒层进行分离,分离后颗粒层放入盛有3%青链霉素PBS的烧杯或培养皿中。
(3)向颗粒细胞层中加入适量胰酶(实验前检测要确定其无污染),放入恒温37℃的CO2培养箱中进行培养,处理15min。
(4)颗粒层处理好后加入PBS终止消化,1800rpm离心5min,弃去上清,加PBS吹洗,1800rpm离心5min,弃去上清。
(6)根据沉淀多少用适量新的培养液把离心管底部的细胞沉淀进行重悬,在24孔板中先加入少量培养液(200ul,第一列四个孔即可),然后每孔加入100uL,150uL,200uL,250uL重悬的细胞于24孔板第一列的四个孔中,用显微镜进行观察,选择适宜的细胞密度进行后续的布板。
(7)每孔中加入适量的细胞悬液,用新的培养液补足充至1000uL,将接种好的板轻轻放入恒温37℃的CO2培养箱进行培养,CO2浓度为5%。
(8) 接种后12h进行换掉部分培养液液,培养24h左右换掉全部培养液,注意换液时用PBS进行清洗,清洗动作轻柔,防止细胞贴壁不紧而导致细胞大量流失。
(9)培养到细胞贴壁85%左右用于后续实验。
2.2.2 Wnt4重组蛋白处理鸡卵泡颗粒细胞
向不同孔的颗粒细胞中分别添加不同剂量的Wnt4重组蛋白(0,5,100,500ng/mL),每个浓度组重复三孔,处理48h后收集细胞。
2.2.3鸡卵泡颗粒细胞的RNA提取
本实验使用Trizol法提取总RNA,详细操作步骤如下:
(1)用PBS清洗两次准备提取RNA的细胞,将1mL Trizol加入培养板的孔中,用移液器吹打,将细胞裂解下来,转移到新的1.5mL的离心管中,置于冰上充分裂解12min左右。4℃,12000rpm,离心90s左右。将适量上清液转移至新的离心管中(避免吸取沉淀)。
(2)向离心管中加入200μL氯仿,用力摇匀,置于室温(15-30℃)静止5min;
(3)4℃ 12000rpm离心15min,管中的液体将分为三层:上层清澈,中间为白色沉淀,下层红色,将上清转移至一个新的离心管中。吸取上清大约为450µL,注意吸上清尽量不要太多,以免吸到蛋白质,污染RNA。
(4)向取得的上清液中加入500µL冰异丙醇,缓慢上下颠倒离心管直至充分混匀,置于冰上10min,沉淀RNA;4℃12000rpm离心10min,弃上清。
(5)向沉淀中加入600µL 70%乙醇,轻微振荡,4℃ 12000rpm离心10min,洗涤RNA,吸出上清。
(6)打开离心管盖,晾干残存的乙醇,约3-5min后加入50μL RNase Free H2O溶解,于-80℃冰箱内分装保存,备用。
(7)RNA质量及浓度检测:电泳检测RNA的质量和浓度,并用紫外分光光度计检测A260/A280、A260/A230的值。
2.2.4RNA反转录后进行qRT-PCR检测
(1)反转录合成cDNA:cDNA的合成按照TakaRa公司反转录试剂盒的操作步骤进行详细步骤如下:
Ⅰ基因组DNA的去除反应
按2-1配成反应体系,置于金属浴中,程序为:42℃ 2min,4℃保存。
Ⅱ反转录反应
按表2-2配成PCR体系,置于PCR仪中进行反转录,程序为:37℃ 15min,85℃ 5s,最后于-20℃保存。
表2-1 DNA去除反应体系组分
| 组分Element | 体积 Volume(μL) |
| gDNA Eraser | 1 |
| 5×DNA Eraser buffer | 2 |
| Total RNA | 1 |
| RNase-free dH2O | Up to 10 |
表2-2反转录反应体系组分
| 组分Element | 体积 Volume(μL) |
| RNase-free dH2O | 4 |
| 5×PrimeScript buffer | 4 |
| RT Primer Mix | 1 |
| PrimeScript RT Enzyme MixⅠ | 1 |
运用Primer Premier 5软件,按照qRT-PCR引物设计原则,设计荧光定量PCR引物,引物由上海生工生物有限公司合成。在荧光定量PCR仪上进行基因mRNA表达水平检测。各cDNA以β-actin为持家基因进行相对定量PCR反应。反应体系见表2-3。荧光定量引物见表2-4。每个样品重复三次。PCR采用两步法,反应条件为:95℃预变性30s;95℃ 5s,60℃ 20s,循环数为 40;熔解曲线:95℃ 1s,65℃ 15s,95℃ 1s。同时将各个样品的cDNA取出等量混池,并以此为模板进行2倍梯度稀释,以持家基因和目的基因特异性引物来进行扩增,建立标准曲线。最后用2-△△Ct的方法算出基因的表达水平[23],确定基因的相对表达量。
表2-3 qRT-PCR反应体系组分
| 组分Element | 体积 Volume(μL) |
| SYBR Premix Ex Taq (2×) | 10.0 |
| cDNA | 1.5 |
| Primer1 | 0.4 |
| Primer2 | 0.4 |
| ddH2O | Up to20 |
表2-4 荧光定量PCR引物
| 基因
Gene | 登录号
Accession Number (GenBank) | 引物序列
Primer Sequence | 产物长度
Product Size |
| FSHR | NM_205079.1 | Forward: 5’ TTCCAGCCTTCCCAAACTA
Reverse:5’ GCCGTAGAATCACACTTTCAG | 258bp |
| StAR | NM_204686 | Forward: 5’ TGCCTGAGCAGCAGGGATTTATCA
Reverse:5’ TGGTTGATGATGGTCTTTGGCAGC | 149bp |
| CYP11A | NM_001001756 | Forward: 5’ ACTTCAAGGGACTGAGCTTTGGGT
Reverse:5’ AGTTCTCCAGGATGTGCATGAGGA | 103bp |
| CTNNB1 | NM_205081.1 | Forward: 5’GTCTCCATTACGGACTCCCA
Reverse:5’CAAACTGCTGTTGCGTTCCACCCAT | 220bp |
| ACTB | K02173.1 | Forward: 5’ TGGATGATGATATTGCTGC
Reverse:5’ ATCTTCTCCATATCATCCC | 253bp |
2.3 数据统计分析
所有数据均用IBS SPSS Statistics 21分析,结果用平均数±标准误表示,不同组之间利用单因素方差ANOVA和邓肯式检验进行,结果均以P<0.05为差异显著。
3.结果
结果如图3-1所示,Wnt4重组蛋白对于StAR和CYP11A1的表达没有显著的影响,但随着Wnt4重组蛋白浓度的增大,StAR和CYP11A1的表达有一定上升的趋势。由此可见,高浓度的Wnt4重组蛋白能够上调StAR和CYP11A1的表达。


图3-1重组蛋白Wnt4对等级卵细胞颗粒中STAR、CYP11A表达的影响
当Wnt4重组蛋白浓度为5ng/mL时,FSHR的表达量出现了显著的上调(图3-2),说明Wnt4重组蛋白对于FSHR的表达具有一定的调控作用。FSHR作为FSH的受体,在卵泡选择的过程中发挥了重要的调控作用。
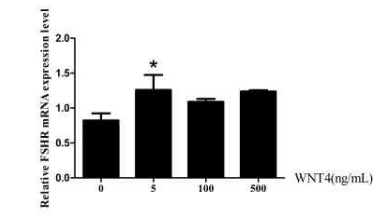
图3-2重组蛋白Wnt4对等级卵细胞颗粒中FSHR表达的影响
当Wnt4重组蛋白浓度为5ng/mL和500ng/mL时,CTNNB1的表达量出现了显著的上调(图3-3),说明Wnt4重组蛋白对于CTNNB1的表达具有一定的调控作用。CTNNB1作为Wnt信号通路的关键成员,对于Wnt信号通路的作用发挥起到了至关重要的作用,也能够在卵泡选择的过程中发挥一定的调控作用。
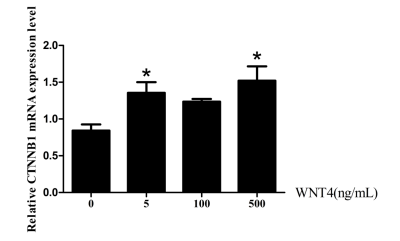
图3-3重组蛋白Wnt4对等级卵细胞颗粒中CTNNB1表达的影响
这些结果说明,Wnt4重组蛋白能够促进相关基因的表达,从而对于等级颗粒细胞的分化和卵泡发育有一定的促进作用,具体的作用机制还需要进一步的研究。
参考文献
[1].王玉玲,胡金朝.我国家禽品种多样性及保护[J].中国动物学会成立65周年年会,1994.
[2].张金壁.类固醇激素合成酶及相关受体表达与猪卵泡闭锁过程的关系研究[J].南京农业大学学位论文,2010.
[3].Sato T, Cheng YN, Wu XC, Zelenitsky DK and Hsiao YF. A Pair of Shelled Eggsinside a Female Dinosaur[J]. Science, 2005, 308–375.
[4].Johnson AL. Ovarian cycles and follicle development in birds[J]. Poult Sci, 2014, 94(4) : 781–785.
[5].Rhee CS,Sen M,Lu D,et al.Wntand frizzled receptors as potential targets for immunotherapy in neck squamous cell carcinomas[J].2002,43(43):6598–6605.
[6].Abramovich D, Irusta G, Parborell F, and Tesone M. Intrabursal injection of vascularendothelial growth factor trap in eCG-treated prepubertal rats inhibits proliferation andincreases apoptosis of follicular cells involving the PI3K/AKT signaling pathway[J]. FertilSteril, 2009, 93(5) : 1369–1377.
[7].Almudevar A, Klebanov LB, Qiu X, Salzman P and Yakovlev AY. Utility of correlationmeasures in analysis of gene expression[J]. NeuroRx, 2006, 3(3) : 384–-395.
[8].Al-Musawi SL, Gladwell RT and Knight PG. Bone morphogenetic protein-6 enhancesgonadotrophin-dependent progesterone and inhibin secretion and expression of mRNAtranscripts encoding gonadotrophin receptors and inhibin/activin subunits in chickengranulosa cells[J]. Reproduction, 2007, 134(2) : 293–306.
[9].Anastas JN and Moon RT. Wntsignalling pathways as therapeutic targets in cancer. NatRev Cancer[J], 2013, 13(1) : 11–26.
[10].Anders S and Huber W. Differential expression analysis for sequence count data. Genome.Biol[J]. 2010, 11(10) : R106.
[11].SchwarzRomondT. Three decades of Wntsignalling[J]. EMBO J, 2012, 2664–2671.
[12].Kawano Y and Kypta R. Secreted antagonists of the Wntsignalling pathway[J]. CellSci, 2003,2627–2634.
[13].Sanchez AM, Vigano P, Quattrone F, Pagliardini L, Papaleo E, Candiani M., Panina-Bordignon P. The Wnt/β-catenin signaling pathway and expression of survivalpromoting genes in luteinized granulosa cells: endometriosis as a paradigm for adysregulated apoptosis pathway[J]. Fertil Steril, 2014, 101(6) : 1688–1696.
[14].AvanesovA, Honeyager SM, Malicki J and Blair SS. The role ofglypicans in Wntinhibitory factor-1 activity and the structural basis of Wif1 effects on Wntand hedgehog signaling[J]. 2012,1785.
LimFT, Ogawa S, Smith AI, and Parhar IS. Proteomics identificationof potential candidates involved in cell proliferation for early stage of brainregeneration in the adult zebrafish[J]. 2017, 10–22. [16].Lucia P, Simona C and Matthias C. An Eye on the WntInhibitory Factor Wif1[J]. 2018, 167–-173.
[17].Ng RC, Matsumaru D, Ho AS, Garcia-Barcelo MM, Yuan ZW, Smith D,et al.Dysregulation of Wntinhibitory factor 1 expressionresulted inaberrant Wnt-beta-catenin signaling and cell death of the cloacaendoderm, and anorectal malformations[J]. Cell Death Differ, 2014, 978–989.
[18].Reguart N, He B, Xu Z, You L, Lee AY, Mazieres J, et al.Cloning andcharacterization of the promoter of human Wntinhibitory factor-1. Biochem[J].2004,229–234.
[19].HuangL, LiMX, WangL,et al. Prognostic value of Wntinhibitory factor-1 expression in hepatocellular carcinoma that is independent of gene methylation[J]. 2011,233–240.
[20].Hu YA, Gu X, Liu J, Yang Y, Yan Y, and Zhao C. Expression patternof Wntinhibitor factor 1(Wif1) during the development in mouse CNS[J]. 2008, 515–522.
[21].KongkhamPN, NorthcottPA, CroulSE, SmithCA, TaylorMD andRutkaJT. The SFRP family of Wntinhibitors function as noveltumor suppressor genes epigenetically silenced in medulloblastoma[J].2010,3017–3024.
[22].RamachandranI, Thavathiru E, RamalingamS,NatarajanG,BenbrookDM,et al. Wntinhibitory factor 1 induces apoptosis andinhibits cervical cancer growth, invasion and angiogenesis in vivo[J]. 2012,2725–2737.
[23].Schmittgen TDand Livak KJ. Analyzing real-time PCR data by the comparative[J]. Nat Protoc, 2008, 3(6) : 1101–1108.
致谢
时光荏苒转瞬之间四年的求学之路即将到达终点,在这两年之中初尝了人生的酸甜苦辣,也在其得到了成长。再次离别之际,我想向所有指导帮助我的老师和陪我共同成长的同学致谢。首先要强烈感谢我的论文指导老师—xxx老师,没有她对我进行了不厌其烦的指导和帮助,无私的为我进行论文的修改和改进,就没有我这篇论文的最终完成。其次,感谢我所有的师长,我从他们身上不仅仅学习到了专业知识,更受到了他们的精神熏陶。最终,我要感谢我的父母和所有的朋友,对我的支持和帮助

1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:1158,如若转载,请注明出处:https://www.447766.cn/chachong/132463.html,