目的
自发性脑出血(sICH)是一系列严重的高起病率、高致死、致残率的脑血管事件。在实验动物模型和细胞水平上,最新研究发现IL-17细胞因子与sICH的发病机制关系密切。此次研究的目的主要是阐明IL-17对sICH患者的在神经功能的临床价值和作用机制。
方法
我们对45例重度脑出血患者以及45例正常健康人的血清IL-17水平进行动态的检测,我们采用建立完善的临床量表对自发性脑出血病人的神经功能损伤程度和神经功能恢复的状况进行详细评估并打分。分别对不同分组的实验小鼠的神经功能损伤进行详细的分析和评价。在此同时,建立细胞再生体系/共培养体系,根据组成成分不同将其分成三组,第一组是NSC培养体系,第二组是NSC+ TCRγδ细胞组,第三组是NSC + TCRγδ细胞+ IL-17中和组。最后我们通过相关实验仪器测试了NSCs(神经干细胞)分化、nestin(巢蛋白)、GFAP(胶质纤维酸性蛋白)和MAP2(成熟神经元特异性标志物)的细胞存活率和不同的生物标志物。
结果
sICH患者血清中的IL-17水平含量与自发性脑出血病人神经功能恢复相互之间呈显著负相关,(p<0.001)。在细胞水平研究中,结果表明在细胞培养/共同培养体系中 ,IL-17中和可能会促进NSC的生存能力,这种NSC的生存能力将受到TCRγδ+共培养体系的影响(p = 0.031) ,TCRγδ细胞共培养的条件下NSCs存活率明显降低(p = 0.023),差异有统计学意义,研究结果认为IL-17中和提高NSC定向分化为星形胶质细胞,而不是定向分化神经元细胞。
结论
IL-17作为一种促进炎症反应的细胞因子,这种细胞因子具有非常强烈的神经细胞毒性作用,这种神经细胞毒性作用显著影响着sICH患者的神经功能恢复状况。可能是由于IL-17促炎因子抑制了NSC向功能神经细胞的分化从而影响病人预后的功能恢复。实验研究得出IL-17中和的功能会削弱IL-17促炎因子抑制作用,IL-17中和不仅可以加强NSCs的定向分化水平,并且可以显著的促进神经功能的恢复。
关键词脑内出血;神经干细胞;白介素17;中和;定向分化
1 前言
自发性脑出血(sICH)是一系列严重的高起病率、高致死、致残率的脑出血疾病的统称,一般由非创伤或者先天疾病因素导致,如高龄、高血压、血清胆固醇水平低、饮酒、颅脑血管畸形和吸烟及医源性用药等甚至不明原因。在中国这种病人的数量约占所有中风病人数量的8%-15%,全世界范围内,在一些发展中国家如非洲或亚洲国家中自发性脑出血患者占中风病人的比重更高,而且往往导致意想不到的坏结果,自发性脑出血患者一个月的死亡率大约占30%-55%,它往往导致较高的致残率及致死率,同时不幸的是超过 30%的存活者遗留神经功能缺损[1,2]。近几十年来,随着医学科学技术的不断进步和发展,为了达到更好的治疗效果,如今医疗技术手段和治疗措施也在不断进步并且高超的技术和手段得到大规模推广和应用,目前全国范围内出现了大型的医疗机构急救中心,目的给了更快捷抢救病人,但是仅能有限的挽救生命或提高病患的生存率,在外科手术治疗方面,比如各种精细操作的开颅手术清除血肿,讲究显微操作小骨窗,起到了极少损坏责任血管或神经,但是经过人类共同的努力,目前仍缺乏有效的治疗方案和措施来降低sICH患者的致残及死亡率或改善脑出血患者的预后。尤其是那些从病理分类中的脑出血急性期幸存下来的患者,虽然生命保住了,可能会引发长期的认知及肢体活动功能障碍,这些预后不良的患者所带来的治疗和术后康复治疗费用,显然会加重家庭和社会的经济负担,与此同时,每况愈下的生活质量也给病人和家属带来心里压力和精神负担[3],因此为了挽救一个家庭,必须研究一种新的方法降低病死率成为医学界攻克的当务之急。
一般情况下,sICH患者起病后患者通常会经历两个病理生理过程:第一个过程是原发性脑损伤,继而发生继发性脑损伤。原发性脑损伤通常在出血后的最初几分钟内发生。因为自发性脑出血患者短时间内扩大的血肿在这个阶段扮演者重要的角色[4],并且血肿量的大小和出血时间直接影响脑出血患者的预后效果包括认知及肢体活动功能,相比较而言,自发性脑出血患者出血时间越短,颅内血肿量越小,往往患者预后越理想,深究其原因,我们认为一定时间内的血肿增加量是导致sICH患者颅内压增高的最重要原因,血肿和颅内压关系可成正相关,随着单位时间内血肿腔的出血量增多,从而使得颅内压随之越高,经研究发现颅内压增高是岛之间sICH患者死亡的重要原因。MacLaughlin 等[5]和 Yuan 等[6]在大量的研究中得出结论均显示颅内压检测仪,在临床治疗中监测患者颅内压,根据颅内压的大小的变化,调整脱水剂的使用量降低患者院内病死率起到了关键性作用。一般常见颅内压增高的因素有多种,其中小脑出血最常见的是由小脑占位性病变所致,随着时间变化,单位时间内血肿出血量的增加导致颅内压增高,造成患者意识上逐渐神智昏迷,最终患者死亡的原因可能是因为枕大孔疝的形成,相比于其他部位,如基底节出血,前者更容易致一定的致残率和致死率[7-8]。从病理学角度分析,以动脉壁发生病理生理改变为基础,一般的血流动力学变化或其它诱因即可引起血管破裂, 此时血液从血管内溢出就引发了临床上常见疾病-自发性脑出血, 自发性脑出血会导致正常脑组织及细胞损伤,形成一定空间的占位效应,当单位时间内出血量迅速增加, 就会导致其穿破了脑实质, 继而引发一系列的病症,如脑室/蛛网膜下腔出血, 若脑出血量增多形成积血可以引起脑室被阻塞或脑表面被压迫,因此脑脊液循环受到影响。同理其占位效应和脑脊液循环受阻导致颅内压随之升高, 等颅高压到一定程度导致脑疝的发生,而固有颅内压的大小、脑出血伴发的损害、出血的速度和量对颅内压增高程度起着关键作用。脑出血时,在血流动力学压力和血肿压力趋于平衡、血管痉挛、 凝血机制三者的共同参与下, 大约30min左右出血会停止, 大约6小时左右血肿不再扩大(头颅CT检查所示)。 颅内压增高除引起导致脑疝的发生外,还可以直接损伤脑细胞以及使脑血管功能发生变化,例如使毛细血管和小静脉受压, 导致小静脉压升高、通透性增加, 最终血浆渗出发生组织水肿。一般在出血6小时左右头颅CT观察可见脑组织水肿, 水肿在3-4天时达到高峰。脑血管功能发生变化亦可发生于小动脉, 此时小动脉出现不同程度的狭窄, 引起脑组织发生缺血缺氧性改变, 从而加重组织损伤。小动脉功能改变所引起的脑组织缺血缺氧性损伤随着颅内压升高以及持续时间延长而逐渐加重, 与病灶距离越远的脑组织损伤越轻。然而,由于脑出血(STICH)的外科手术治疗并未能提供充足并具有说服力的证据来显示出早期手术切除血肿的疗效[9],所以研究者们的目光转移到了对继发性脑损伤的研究上来,以探索自发性脑出血的发病机制和治疗原则及策略为研究重中之重。sICH产生的继发性脑损伤可能的原因是由于脑实质内血液的存在而触发一系列的连锁反应,自发性脑出血后的脑内炎症反应主要表现为各种活化的炎症细胞的积聚。这些反应细胞释放炎症介质,包括趋化因子、细胞因子、蛋白酶和其他促炎分子。进而激活细胞毒性、兴奋性、氧化性和炎性途径等[10-12]。由于血脑屏障(blood-brain barrier, BBB)通常在sICH发病后被中断,在外周循环中的中性粒细胞和各种免疫细胞趁机在被破坏后的血脑屏障中逐渐浸润到大脑组织中去,相应引发脑内各种细胞产生一系列的连锁性炎症反应。因此,自发性脑出血患者局部的一系列的炎症反应通常起关键作用,并与sICH的病理过程和患者功能恢复有密切联系[13,14]。
在sICH患者所有治疗方案中,在有时效性的临床前试验中,证实NSCs移植具有良好的治疗预后效果,试验证实NSCs移植可以有效地促进神经干细胞的发育, 从而改善脑卒中动物的神经功能缺陷,神经干细胞移植,它的特点是具有较强的自我更新和多向分化的能力,因此在sICH患者中把NSCs移植作为一种潜在的治疗策略[15]。然而,NSCs的作用机制,特别是NSCs如何定向分化为某些特定的功能神经细胞,截止到目前临床实验验证中仍未得到明确的答案,NSCs如何分化的机制一直是临床应用中需要及时解决的关键问题。由于sICH患者局部性和全身性发生着复杂的病理生理性及综合病变的改变,炎症相关的微环境与NSCs的分化可能相互作用,进而进一步影响患者神经系统功能的恢复[16,17]。
一些白介素家族的细胞因子,包括IL-1β, IL-6, IL-8 and IL-10,我们对这些细胞因子的发病机制的研究进行了深入的探索[18]。最近研究表明IL-17,一种从TCRγδ细胞分泌的细胞因子,研究结果证实IL-17这种细胞因子在动物实验模型与中风的发病机理密切的关联[19、20]。除此之外,基于当前的实验研究结果证明IL-17中和在应用于某些中枢神经系统疾病有显著的治疗效果[17]。然而,IL-17在sICH发病机制中神经功能恢复中的临床价值和作用机制一直是尚待解决的谜团。在本研究中,我们将首先探讨IL-17在sICH患者中的临床价值,同时也继续深入探讨IL-17在体内发病机制及体外神经发生中的潜在的影响及作用。
研究方案
本课题的研究分为两部分
第一部分:首先采集重度自发性脑出血患者以及正常健康人血清标本,然后对人体血清中IL-17水平进行动态的检测,通过检测自发性脑出血以及健康人群中血清IL-17含量变化来比较两者之间的差异性,并且采用相对应的临床量表(包括GCS评分量表及GOS评分量表)对sICH病人的神经功能损伤程度以及神经功能恢复的状况进行详细评估。
第二部分:从细胞水平,建立细胞模型分组,第一组是NSCs培养体系,第二组是NSCs+TCRγδ细胞培养体系,第三组是nsc + TCRγδ细胞+ IL-17中和培养体系,检测时间分别在24h-72h内每隔24h测一次,运用蛋白免疫印迹技术,ELISA检测和CCK-8检测TCRγδ细胞及IL-17单抗于神经干细胞共培养对IL-17表达影响,来探讨血清IL-17细胞因子水平与临床病理特征之间的关联性,
2 材料与方法
2.1 临床资料
我们收集样本的来源是从2017年1月至2018年3月份期间,从安徽医科大学第一附属医院神经外科并且所有患者均在症状出现后4小时内就诊于我院急诊科治疗的45例重度自发性脑出血患者,以及45例地域、年龄、性别相匹配的健康对照组,所有对照组都是从我院门诊体检中心筛选的健康人群,实验组及对照组之间的种族、地域、年龄、性别等人口统计学资料之间无明显统计学差异,所有这些患者均符合各种脑血管病诊断关键要点所述的ICH诊断标准。纳入标准如下:1、头颅CT(computed tomography)或者头颅MRI(Magnetic Resonance Imaging)提示颅内出血;2、至少3名5年以上从医经历的副主任医师诊断为出血性脑卒中;3、收缩压<180 mmHg,舒张压<110 mmHg。
自发性脑出血的排除标准:1)因其他原因(经过全脑血管造影或脑血管成像检查等筛选化为不明原因)引起的脑内出血,2)由车祸或高处坠落伤导致的外伤性颅内血肿等外伤性的脑内出血性疾病;3)患者本身有严重基础疾病、比如严重慢性心肺功能疾病,循环和呼吸功能不稳定的患者24h内导致死亡、肾衰竭在透析过程的患者e、特发性血小板减少症患者、血友病患者等;4)由长期服用影响血凝机制的药物导致的脑内出血;比如长期服用三七、华法令房颤患者、长期服用阿司匹林凝血机制不良患者;5)患者既往有恶性肿瘤或良性肿瘤,行颅内肿瘤切除术后或自发性脑出血术后引发术区再次出血的患者等;6)患者有神经系统性疾病;
健康组剔除标准;1)患有严重的免疫系统性疾病、恶性肿瘤、严重意识障碍等。2)患者有中枢神经系统和外周神经系统的疾病等。
纳入原则:调查人员通过设计流行病学调查表进行流行病学调查,并详细记录所有纳入实验的患者样本临床流行病学资料,纳入的信息一般包含(性别及年龄)家族史、食物习惯、抽烟情况、长期高血脂情况、是否有饮酒情况、用药史(抗凝药物、抗血小板聚集药物)、长期的血压控制不佳、长期血糖控制不佳、颅内出血血肿大小、部位(位于幕上还是幕下)、血肿边缘距离皮层的多少、血肿压迫各脑室及脑池程度、中线移位程度、脑出血是否破入脑室、是否形成脑积水等。
全部流行病学资料统计是由专职培训人员进行记录及汇总,目的保证资料的可靠性及完整性,参与本研究的自发性脑出血患者均提前告知其被纳入实验当中,所有患者和对照组均获得患者或家属知情同意。所有涉及人类参与研究的临床验证执行的程序均经中国安徽医科大学临床研究伦理委员会批准,且符合机构和国家的道德标准,并且按照《赫尔辛基原则宣言》开展进行的。
纳入人群处理事项:这些样本收集的时间是从2017年1月至2018年3月份。并且所有患者均在症状出现后4小时内入院。然后在入院时(第1天、第3天、第7天和第14天)通过连续收集患者静脉血清样本。按照同样的收集方法,我们从对照组中收集健康人群静脉血清样本进行健康检查。所有血清样本需要通过专业操作者操作离心机分层后抽取上层淡黄色血清样本放入抗凝试管中,特别注意,所有经分离处理后患者的血清样本均储存于-80℃。为了避免血清蛋白含量被降解,因此采集后所有患者血清样本尽量防止反复冻融,与此同时,我们采用格拉斯哥昏迷评分(GCS)对每位患者入院时的神经损伤程度进行评估并打分,入院后第14天的患者恢复情况我们采用格拉斯哥预后评分(GOS)对进行评估打分。两名以上10年以上从医经历的临床副主任医师采用盲法的方案对患者进行评估打分并取平均值。
人血清采集处理方法:实验采用的真空采血管是含有EDTA抗凝剂,患者前臂静脉通常被选为穿刺部位进行采集样本,一般抽取前臂浅静脉血标本容量为5-10ml左右,采集完毕后小心轻放抗凝管,避免红细胞破裂溶解,用ELISA试验方法来检测人血清和不同共培养细胞组的IL-17细胞因子的水平。
2.2 细胞水平实验方法:
试剂选择
除另有说明外,所有试验当中所需的试剂均为商业获得并严格按照规定使用。小鼠NSC系(NE-4C)来自中国科学院细胞库(上海,中国)。抗体老鼠 IL-17A中和抗体都是从BD生物科学公司购买(XCA)。抗小鼠GFAP抗体、抗小鼠MAP2抗体和抗小鼠nestin一抗Western Blotting (蛋白印迹法)检测的主要抗体都是统一购自伊莱瑞特公司(武汉,中国)。用于人类和小鼠IL-17A检测的酶联免疫吸附试验(ELISA)试剂盒购自瑞博奥公司(GA, X)。CCK-8试剂盒购自X西格玛公司。
共培养体系
1.细胞模型分组
第一组是NSCs培养体系,第二组是NSCs+TCRγδ细胞培养体系,第三组是nsc + TCRγδ细胞+ IL-17中和培养体系,检测时间分别在24h-72h内每隔24h测一次
第一组是NSCs培养体系
小鼠神经干细胞NE-4C细胞系的常规培养
首先强调的是整个无菌操作都应该在酒精灯的周围,将细胞培养将小鼠神经干细胞冷冻后的NE-4C细胞立即在37℃水浴中进行预处理,目的是将其缓慢解冻。然后将NE-4C细胞放置在容量为2ml,含有1%抗青霉素-链霉素双抗体,1%谷氨酰胺,1%非必需氨基酸,10%热灭活胎牛血清(FBS)的杜尔贝克改良的eagle培养基(DMEM),置于37℃、5% CO2、时间控制在20 分钟,相对湿度为95%的适宜条件下的细胞培养箱中培养,分别经过将细胞复苏及换液后,传代培养于无菌培养瓶中(比例为1:2或1:3),加入培养基后继续培养或实验。
第二组是NSCs+TCRγδ细胞培养体系,
细胞样本的制备:工作人员将生长状态良好的小鼠NE-4C细胞接种于多聚赖氨酸包被的细胞培养板,待NE-4C细胞完全贴壁后;正常对照组常规培养,实验组按照NE-4C: γδT =1:5的比例加入γδT细胞共培养24h。其中抗IL-17单抗组同时加入2ug/ml IL-17抗体共同孵育24h。收集细胞上清液进行后续ELISA 检测
第三组是nsc + TCRγδ细胞+ IL-17中和培养体系,
用浓度为2μg /ml的中和抗il -17单克隆抗体添加到nsc + TCRγδ细胞培养体系中,然后将这些细胞在培养基中共培养24小时。
2.ELISA检测TCRγδ细胞及IL-17单抗于神经干细胞共培养对IL-17表达影响
2.1主要实验仪器及试剂:胰蛋白酶、MEM培养基、FBS、PBS、青霉素、链霉素电热压力蒸汽消毒器(YXQ-LS-100A)、超净工作台 、二氧化碳培养箱(MCO-15AC-SC)、白细胞介素17(IL-17)ELISA 试剂盒、高速台式离心机(TG16-WS) 、细胞培养瓶及细胞培养板(Costar)、细胞计数板光学显微(OLYMPUS CK-40) 、倒置显微镜(OLYMPUS)
2.2本实验原理:依据双抗体夹心法
应用酶联免疫分析技术检测小鼠样本中白介素-17(IL-17)的水平。其中96孔板含有纯化的小鼠IL-17抗体,此为本试剂盒的固相抗体,依次往含有IL-17单抗的孔内加入适宜的小鼠IL-17,最后结合被HRP标记的IL-17抗体,形成了抗体-抗原-酶标抗体复合物,也就是我们常说的“双抗体夹心”法的由来。酶标板加样本反应后,经洗板后加显色底物TMB。显色底物TMB经酶(HRP)的催化作用后可发生颜色反应,相应的转化成蓝色,并在酸性条件下转化成黄色的最终色。根据颜色的深浅和样品中的小鼠白细胞介素-17呈正相关。用酶标仪测定酶标孔各孔OD450吸光度值,根据标准曲线所得公式计算出样本的中IL-17的水平。
2.3 ELISA检测步骤
2.3.1样本收集处理
细胞分组培养后,用吸管收集细胞上清液于离心管中(无菌),将离心管放入离心机中离心20min(转速:3000转/分),吸管吸取离心后的上清液以备后续ELISA检测。
2.3.2 ELISA检测
1.标准品的稀释与加样:在酶标板上选择好标准品孔(10孔),在第1、第2孔中均加入标准品100μl、标准品稀释液一半浓度,移液枪吹打混匀。然后从第1孔、第2孔中已经混匀的液体用移液枪分别取100μl,加入到第3孔和第4孔中同样的进行吹打混匀;于第3孔和第4孔中每孔弃去50μl孔内液体,然后再从3、4孔各吸取50μl液体分别加到第5与第6孔内,参照以前步骤混匀后以此类推,最后从第九第十孔中各取50μl弃掉。
2. 加样:先设立空白孔(调零用),其中空白对照孔既不加待测样本也不加标准品,然后于每个样品孔内加入相应的样本。将50μl待测样本(每孔)加入到酶标包被板中的待测样品孔中。加样时要注意将样品加于酶标板中孔的底部,轻轻晃动或用移液枪进行吹打混匀,切忌触到酶标板孔壁。
3. 温育:用封板膜封板后,在37℃温度下进行温育三十分钟反应。
4. 配液:按说明书推荐比例,用蒸馏水稀释20×浓缩洗涤液,备用。
5. 洗涤:温育完成后将封板膜小心揭去,弃去酶标板内液体,轻轻甩干后,每孔加满稀释好的洗涤液洗板,静置30s左右弃去洗涤液,重复操作洗板5次,最后轻轻拍干酶标板孔内的液体。
6. 加酶:96孔酶标板中每孔加入50μl酶标试剂(空白孔不加)。
7. 再次进行上述温育及洗涤步骤。
9. 显色:每孔均加入显色剂A&B 50μl:50μl,通过轻微震荡混合均匀后在37℃条件下避光反应15分钟进行显色。
10. 终止:将50μl终止液加入各孔内使反应停止(此时蓝色立即变为黄色)。
11. 测定:以空白孔为基准进行调零后,在酶标仪上读出各孔在450nm波长处的吸光度值(OD450值)。注意加入终止液后15分钟以内应尽快上酶标仪进行测量OD450值。
2.4计算方法
在坐标纸上画出横坐标(标准品的浓度)和纵坐标(标准品OD值),得出所需的标准曲线,以此为据算出标准曲线的直线回归方程式,将酶标仪所测得的各孔样本OD值代入方程式,计算出所求样本的浓度(须乘以稀释倍数)。
3.用CCK-8检测TCRγδ细胞及IL-17单抗与皮层神经元共培养对神经干
细胞存活影响
3.1基本原理
CCK-8试剂盒(Cell Counting Kit)最重要的材料是WST-8(化学名:2-(2-甲氧基-4-硝苯基)-3-(4-硝苯基)-5-(2,4-二磺基苯)-2H-四唑单钠盐),此检测方法应用非常广泛,例如可以用于快速检测肿瘤细胞药敏试验、细胞增殖、细胞毒性、药物筛选以及细胞因子活性检测等。本试剂盒工作原理可概括为在电子耦合剂环境下,线粒体内的脱氢酶促使WST-8经过还原反应生成橙黄色并且具有水溶性的产物。此反应产生的橙黄色深浅也代表着细胞的增殖程度,并与之呈正比,但是与所反映的细胞毒性成反比。细胞数量可以用酶标仪测定的OD450值来间接表示。
物品准备:
首先研究人员备好实验相关器材及用具:电热压力蒸汽消毒器、超净工作台、二氧化碳培养箱、低速台式离心机、多功能酶标仪、移液器细胞计数板、MEM培养基胎牛血清、青霉素-链霉素溶液(100X)、胰蛋白酶、PBS、CCK-8试剂盒
3.2操作步骤
1 细胞培养
小鼠神经干细胞NE-4C培养于MEM培养基中(含10%优质胎牛血清和1%青霉素-链霉素双抗,1%Glutamax,1%Non-essential Amino Acids)进行培养,培养条件: 37℃、5%CO2、相对湿度为100%。经过传代后留待CCK-8实验检测分析。
2 CCK-8检测
于96孔板中接种100 μL已经计数调整浓度(调整浓度为1×105/mL)的样本,每个稀释倍数的细胞样本设置三个复孔以减少误差。在细胞培养箱中(37℃,5% CO2)培养96孔板中细胞样本,设置的培养时间分别为24、48、72小时三个时间点,每孔加入10μL CCK-8工作溶液,切忌往96孔板加液时不要产生气泡,否则严重影响实验结果。并置于培养箱内孵育1-4h。
3活力计算
| 测定目标 | 计算公式 |
| 细胞增殖/毒性活力(%) | [A(实验组)-A(空白组)]/[A(对照组)-A(空白组)] ×100% |
| 细胞抑制率(%) | [A(对照组)-A(实验组)]/A(对照组)=1-细胞活力 |
注:①A(实验组):具有经过处理的细胞、CCK溶液的吸光度值;
②A(空白组):具有培养基和CCK溶液而没有细胞的孔的吸光度值;
③A(对照组):具有未经过处理的细胞、CCK溶液的吸光度值。
4.免疫印迹分析检测γδTT细胞及IL-17单抗与皮层神经元共培养对神经干细胞分化影响
蛋白免疫印迹( Western Blot) 的具体原理是将组织细胞所有的蛋白分子大小不同,通过电泳可以将不同大小的蛋白分离开后,然后将凝胶上电泳分离的蛋白通过电流在转膜液中转移到NC膜或者PVDF 膜上,由于抗原抗体可以特异性结合,用特异性抗体的方式就可以识别检测某特定抗原(即目的蛋白)。Western Blot作为一种蛋白质检测分析的实验技术,抗体的作用可以理解为“探针”,通过“探针”检测识别特异的蛋白质。而最终识别结果是通过用辣根过氧化物酶标记的二抗来进行显色识别。抗原转移到固相载体(NC膜或者PVDF 膜)上形成蛋白质或多肽,与其特异的一抗发生反应形成抗原抗体复合物,接下来一抗再与辣根过氧化物酶或者也可能是同位素标记的二抗发生免疫反应结合,最终经过底物显色或放射自显影反应来检测电泳分离的特异性目的基因所表达的目的蛋白。目前在抗体活性的研究检测、蛋白水平的表达、以及某些疾病的早期诊断等领域均可见到Western Blot的应用。
WB操作步骤
1.总蛋白提取
1.1细胞裂解
首先进行总蛋白质的提取,在提取过程中可以先进行细胞裂解 具体步骤是将裂解后的细胞将贴到壁细胞,然后丢弃培养基,用PBS进行第一次清洗。再者针对悬浮细胞,具体操作是先收集所需细胞置于低温离心机进行离心后,用PBS缓冲液再次进行清洗,向细胞中加入RIPA裂解液(比例:每106个细胞加0.1 mL RIPA buffer),同时要特别注意的是使得裂解液和细胞充分的接触,可以在冰上动作轻柔的用枪头吹打细胞液使得裂解液和细胞充分的接触达到细胞充分裂解的目的。再轻轻倾斜培养皿使裂解产物顺利的流向另外一边,然后将裂解的细胞产物转移至离心管(1.5 mL)前需通过培养皿轻轻倾斜促进裂解产物顺利的流向另外一边以便进行转移,维持在30秒,以转速12000rpm进行大幅度的振荡,在4 ºC低温离心机离心15min,转移离心后离心管内液体的上清液,以备进行后续的Western Blot操作,
1.2蛋白浓度测定(BCA测蛋白浓度)以及工作液的配制
进行电泳前需先按照实验需求进行BCA测蛋白浓度测定,测定前首先配制好BCA法工作液(比例Reagent A:BCA Reagent B = 100:1)。用BCA来测蛋白浓度,至关重要的一步是进行BSA 标准品的配制以求得标准曲线。
(1)稀释BSA标准品溶液:取120μL BSA 标准品 (2mg/mL),加入1080 μL稀释液后充分混合配置成标准品稀释液(0.2 mg/mL)。
(2)配制各标准孔内标准品溶液(见下表)。采用PBS缓冲液稀释 BSA 标准品和待检测的样本。
| 0.2mg/mL BSA Standard(uL) | 稀释液(uL) | BSA终浓度(ug/mL) |
| 400 | 0 | 200 |
| 300 | 100 | 150 |
| 200 | 200 | 100 |
| 100 | 300 | 50 |
| 40 | 360 | 20 |
| 20 | 380 | 10 |
| 10 | 390 | 5 |
| 0 | 400 | 0 |
1.3BSA 标准曲线的制备
同理实验至关重要的一步是BSA 标准曲线的制备,具体步骤为,在微孔板中分别加入已经稀释的BSA标准品溶液100 μL/孔,采取每个浓度都要设置二个复孔以减少误差,然后同样的在微孔板中上述加入标准品的孔内分别加入100uL已配置好的BCA工作液,立即进行混匀并且在37℃水浴槽中反应60分钟,时间到后取出微孔板冷却至室温,选定波长在562nm处并应用分光光度计测定时微孔板各孔的吸光度值。注意关于测量数值,可以选择使用1 mL比色皿,用双蒸水调零。检测完毕所有样品尽量控制在20min内,得出数值后,将分光光度计测得的各孔BSA标准品溶液的OD562值减去空白孔OD562值进行调零后,绘制出标准品的标准曲线。待测细胞样本与BSA 标准品溶液同时进行测定OD562值,最后依据所得出的标准曲线推算出待测细胞样本总的蛋白浓度。
1.4 SDS-PAGE电泳
再进行最关键的一部分SDS-PAGE垂直电泳,详细步骤如下:
选择洁净、干燥且与制胶架配套的长短玻璃板,看好正反面对齐后按照正确方向插入制胶架中卡紧,然后垂直卡在制胶架子上准备灌胶(预先加入生理盐水确认制胶架契合严密不漏胶),按比例配制出8%,10%分离胶,加入TEMED后立即轻柔摇匀混合,马上将配好的分离胶灌入两块玻璃板间,加至适量高度(在绿线附近即可),注意动作缓慢轻柔避免产生气泡及胶漏出,最后加入双蒸水起到隔绝空气加快胶凝固以及压胶的作用,说明胶已凝成功的标志是水和胶之间有一条折射线,水和胶之间有一条折射线后要再等3分钟左右,就可倾斜制胶架倒去胶上的双蒸水并用滤纸将双蒸水尽量吸干,然后再根据比例配制出浓度为5%的浓缩胶,加入TEMED后轻柔摇匀混合即可灌胶,马上将将梳子插进灌好的浓缩胶中,待浓缩胶凝固后,取出含胶的玻璃板置于电泳夹中卡紧(注意玻璃板正反顺序)后放入电泳槽内。取出按BCA法计算所得蛋白浓度配制的上样样品,与5×SDS上样缓冲液配比后混合并在沸水中高温煮5min使蛋白变性。按每泳道等量蛋白上样原则进行上样后,向电泳槽内加入适量电泳液,正确连接电源后选择浓缩胶80V压缩条带,等溴酚蓝指示条带压缩成一条直线后,转换电压至120V将目的蛋白条带跑开,待溴酚蓝跑至胶板底部刚好没有跑出时,可关闭电源,取出夹子将夹子打开使黑的一面保持水平向下,并在黑色夹子一面放上海绵垫→滤纸→含有目的蛋白的凝胶→已被甲醇激活的PVDF膜→滤纸→海绵垫,然后将槽内的电泳液倒去并改为加入转膜液,正确连接电源后选择恒流200mA,转膜约1~3小时终止。越大的蛋白转膜时所用的时间也越长,这是根据蛋白分子量的大小来判定的,可控时间范围内目的蛋白都则会转移到PVDF膜上。2.接下来取下夹子取出膜,并用剪刀或铅笔标记住正反面,用配好的TBST溶液摇床上清洗1分钟,然后配制5%脱脂牛奶作为封闭液室温封闭PVDF膜2小时左右。一抗应用5%脱脂牛奶封闭液进行稀释成一定的浓度(比例大约稀释到1:500),然后将PVDF膜与稀释好的相应一抗反应,条件为4℃孵育过夜,二抗反应选择室温孵育1.5h左右。用TBST洗3次×5min/次。用封闭液将二抗按一定比例稀释(比例大约稀释到1:1000),二抗反应选择室温孵育1.5h左右。再重复用TBST在摇床上清洗4次×5min/次,将ECL发光液按A液:B液等比例配比并混匀,均匀滴加在整片PVDF膜上反应2min,最后在曝光仪上进行曝光检测。利用Tanon 1600R成像系统(Tanon Technology Co. Ltd, Shanghai, China)对蛋白条带进行可视化捕获。每个波段的光密度是量化使用ImageJ软件(贝塞斯达国立卫生研究院,医学博士,X)和归一化β-actin强度。
2.3 统计学分析
选择SPSS 19.0对实验结果进行统计学分析。本研究所有列举的数据均以均数±标准差(x ± s)表示,采用Kruskal-Wallis检验进行分析。采用卡方检验进行二元数据分析。以P<0.05为差异有统计学意义。
3 结果
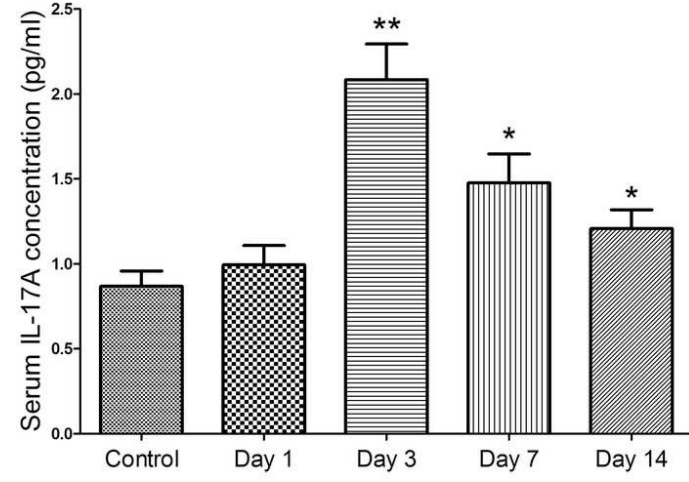

图1所示 sICH患者血清IL-17细胞因子水平的动态变化
Figure 1. Dynamic changes of serum IL-17 level in sICH patients
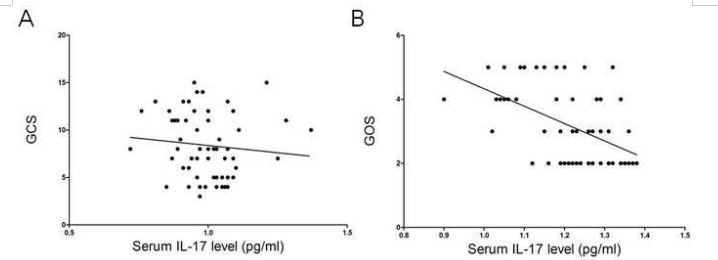
图2 入院时GCS和第14天GOS的血清IL-17水平的相关性分析
Figure 2. Correlation analysis of serum IL-17 level with GCS at admission and with GOS at day 14
3.1 sICH患者血清IL-17水平升高并神经功能恢复的关系
SICH患者血清中的IL-17水平在第3天明显升高,IL-17水平含量与神经损伤和功能恢复密切相关,我们为了探讨IL-17与sICH病理过程的临床价值以及关系,因此在不同时间点动态检测人血清IL-17水平。结果显示,与健康对照组相比,SICH患者血清IL-17水平明显升高,在第3天达到峰值,随后下降到一个相对较高的水平(如图1所示)。与此同时,我们分别使用GCS和GOS对血清中的IL-17水平与神经功能损伤及恢复进行相关性分析。分析结果表明患者入院时血清中的IL-17水平与GCS显示负相关性(相关效率r2=-0.211,p=0.105),而入院第14天患者血清中的IL-17水平与GOS呈显著负相关(相关效率r2=-0.498,p<0.001)。(如图2所示)
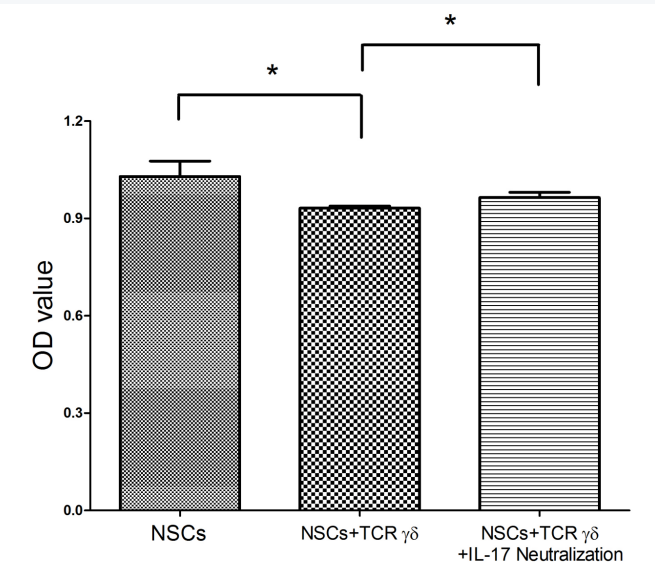
图3不同细胞培养/共培养组间IL-17浓度检测。
Figure 3 Detection of IL-17 concentrations in different cell culture/co-culture groups
图4 不同细胞培养/共培养组间的细胞活力测试。
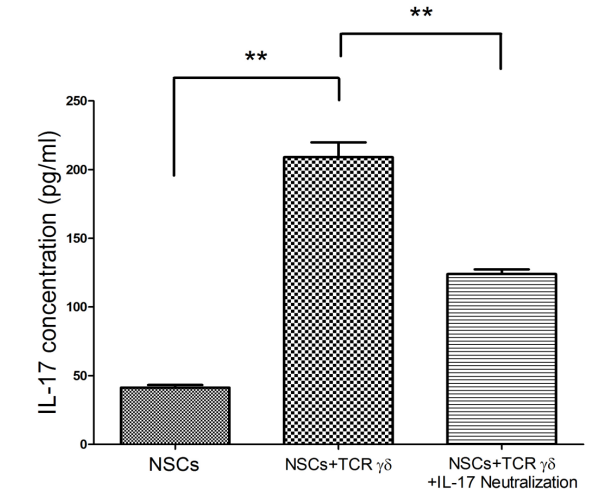
Fig.4 Cell viability test between different cell culture / coculture groups
3.2 IL-17中和可以消除IL-17对NSCs生存活力的不良影响
首先,我们测试了抗IL-17单克隆抗体是否能有效中和功能性IL-17浓度,在细胞培养系统中IL-17这种细胞因子主要是由TCRγδ细胞分泌的,我们通过ELISA这种染色方法来检测IL-17细胞因子的水平含量(图3所示)。最终根据实验研究结果表明;抗IL-17单克隆抗体可以有效的抑制TCRγδ细胞分泌IL-17这种细胞因子。然后我们研究人员用CCK-8这种试剂来检测这三个实验小组间72小时内的NSCs存活率。根据结果显示;TCRγδ细胞共培养的条件下NSCs存活率明显降低(p = 0.023),差异有统计学意义,换句话说,TCRγδ细胞可以分泌IL-17细胞因子,而IL-17细胞因子可以降低NSCs生存率。
然而,相比,nsc + TCRγδ共培养细胞组中,我们得出的结论是;IL-17中和可能改善nsc生存能力,并且可以增加了nsc生存能力及数量,(p = 0.031),差异有统计学意义。(图4所示)。
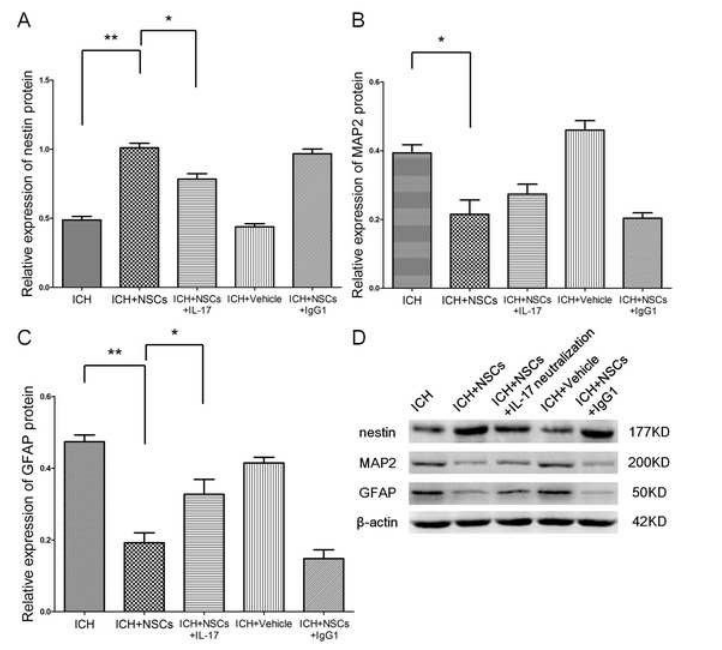
图5 Western blotting凝胶图像(d)显示nestin (a)、GFAP (b)和MAP2 (c)在不同细胞培养/共培养组中的表达。
Figure 5 Western blotting gel image (d) shows the expression of nestin (a), GFAP (b) and MAP2(c) in different cell culture/coculture groups
3.3 TCRγδ细胞分泌IL-17和IL-17中和可以促进NSCs细胞分化作用
本节主要介绍通过Nestin、MAP2和GFAP这三种蛋白,来评估IL-17细胞因子和IL-17中和对这三组实验中的NSCs分化的影响。根据实验结果我们得出;相比对照组(nsc细胞培养组)来说,nsc + TCRγδ共培养细胞组中的巢蛋白(Nestin)的表达是明显呈上升趋势,而且根据实验所得数据中得出p < 0.001,差异有统计学意义。然而,相比nsc + TCRγδ培养细胞群中来说,巢蛋白在nsc + TCRγδ+ IL-17中和培养细胞组的表达是明显呈下降趋势,(p = 0.002),差异具有统计学意义。研究结果显示,我们不难发现相比对照组来讲,GFAP这种蛋白在nsc + TCRγδ共培养细胞组中表达是明显呈下降趋势(p = 0.003),然而,当实验组中加入IL-17中和后,GFAP这种蛋白的表达是呈上升趋势的,(p = 0.015,差异具有统计学意义,也就是说IL-17中和可以有效的抑制TCRγδ细胞分泌IL-17,当IL-17减少时,炎症细胞因子跟着下降,反而相对应的GFAP蛋白表达明显上升。我们根据MAP2蛋白表达的所产生的效果显示,我们实验研究发现在nsc + TCRγδ共培养细胞组中,MAP2蛋白表达明显呈下降趋势。然而,我们并没有发现的是在nsc + TCRγδ培养细胞组和nsc + TCRγδ+ IL-17中和培养细胞组,MAP2蛋白在2组实验结果之间的表达有统计学差异(p = 0.098),也就是说MAP2蛋白在nsc + TCRγδ培养细胞组和nsc + TCRγδ+ IL-17中和培养细胞组表达是没有区别的(如图5所示)。
4 讨论
综上所述,经过查阅大量现有文献可知,NSCs是一类具有自我更新替代能力并具有多能性的细胞群,同时NSCs具有可以分化为若干细胞系的特性,比如说较为常见的是神经元、星形胶质细胞和少突胶质细胞等。正常情况下,NSCs通常位于大脑的脑室下区(SVZ)和亚颗粒区(SGZ),而且NSCs在分化特性中并不活跃,但是在人体大脑发育的每个阶段中均发挥举足轻重的作用。然而根据近来研究结果的说法表明,非活性NSCs可以在大脑受到损伤后很快被激活,被激活之后的NSCs可以参与到大脑细胞的修复过程中去。特别的是,对于大脑受到损伤后发生的整个病理反应过程而言,被激活后的NSCs参与的自我修复作用往往是短暂的、不充分的[22]。因此,近几十年来,世界范围内的研究人员对颅脑损伤中,包括外伤性脑出血以及自发性脑出血,NSCs的研究数量逐年增加,研究人员对脑损伤中NSCs的研究目的;主要是想发现激活内源性NSCs和移植外源性NSCs在促进神经功能损伤后修复过程中的潜在治疗策略[23,24]。同样在这个阶段,我们研究人员所关注的是,神经炎症在ICH微环境下的继发性脑损伤过程中起着基础性作用。同样实验结果中我们也发现的是多种抗炎细胞因子、促炎细胞因子都参与了神经功能损伤和神经功能修复的过程中,而且抗炎细胞因子、促炎细胞因子在NSCs移植的分化和增殖过程中发挥全面性的相互作用[25]。
最近研究报道的,IL-17的结构特点IL-17既是Th17细胞分泌的特征性细胞因子,同时又是Th17细胞的极为重要的效应细胞因子。到目前为止已有命名为6个IL-17家族成员(IL-17A、IL-17B IL-17C, IL-17D, IL-17E和IL-17F)和5种IL-17受体(IL-17RA, IL – 17 rb、IL-17RC IL-17RD和海基会).IL-17A(即IL-17)与IL-17F在结构上极度的相似,其同源性达到50%,人的IL-17和IL-17F基因均附着在染色体的6 p12位点,其中IL-17的长度是4292个基点,是由155个氨基酸残基依靠二硫键的结合形成的,其相对分子质量为(30 ~ 35)×103的同源二聚体,实验得出,Th17细胞产生的IL-17和IL-17F,主要是以IL-17 / IL-17F异二聚体的形式存在[26],IL-17和IL-17F可与上皮细胞,内皮细胞,以及巨噬细胞上的IL-17RA和IL-17RC相结合,诱发表达细胞因子(IL-1β肿瘤坏死因子,il – 6, GM – CSF)、趋化因子(处于受控,CXCL8 CXCL10)以及金属蛋白酶,从而发挥促炎生物学效应在炎性部位上[27]。其同样也被称为IL-17A细胞因子,IL-17A是IL-17家族的主要的细胞因子的其中一份子,由T (Th17)细胞的一个亚群产生(Huber et al., 2013; Costa et al.,2012; Coccia et al., 2012)。尽管IL-17A单独的作用效果是有限的,目前已经研究证实的是,IL-17A与IL-1β,IL-22以及体内其他细胞因子之间相互合作,(Adamik et al., 2013; Abe et al., 2012; Dungan andMills, 2011)。特别是IL-17A会诱发促炎性反应,通过诱导许多其他细胞因子和趋化因子的反应(Newcomb et al.2012;Rocha等人),这种细胞因子是在先天免疫和获得性免疫之间的一种密不可分的联系,据报道IL-17A细胞因子在神经炎症反应中发挥的作用中是一把双刃剑,在某些反应中发挥有利作用,然而在另外某些反应中发挥着不利的作用[28]。然而,迄今为止,世界范围内关于IL-17这种细胞因子在sICH的诊断方略和预后康复治疗方面的临床价值的研究仍比较少。一项对欧洲多中心的研究中我们不难发现,无论是ICH还是脑缺血超急性期,循环IL-17细胞因子的水平可能不是区分卒中模拟物与真实卒中的候选生物标志物[29。在现有的实验研究结果表明,相比一些在ICH早期即有反应的许多生物标志物来说,比如IL-4、IL-10和IL-6等其他细胞因子来讲,我们清楚的检测到血清IL-17细胞因子在ICH患者入院发病后的第3天水平含量是明显上升的[30,31]。同时,我们在ICH患者入院时通过GCS评估并未发现血清IL-17细胞因子的水平与患者的神经功能损伤之间有相关关系,根据评估结果提示血清IL-17在ICH患者的病理过程中是一种比较缓慢的病理反应过程,换句话说ICH患者最开始病情发展时,这个血清IL-17并没有参与其中。同时,我们通过评估患者GOS评分中发现,血清IL-17细胞因子在第14天水平含量升高,血清IL-17细胞因子水平含量升高的ICH患者的恢复比较慢,预后也比较差,提示IL-17细胞因子在脑出血发病过程中具有神经毒性作用,也就是说血清IL-17细胞因子抑制神经功能恢复,这种情况持续进展的话可能对疾病的预后产生不良影响。
TCRγδ细胞作为主要的IL-17捐赠者,也就是说TCRγδ细胞分泌IL-17这种细胞因子,而且我们通过文献查不难发现是TCRγδ细胞通过向周围的骨髓细胞(比如中性粒细胞和单核细胞)产生各种趋化信号,这种信号可以驱动特殊的细胞分泌特殊的细胞因子,比如说TCRγδ细胞分泌细胞因子中,其中包括IL-17在内的,这些细胞因子在分泌之后,作用于靶细胞中有神经毒性一般在神经损伤后,导致缺血性脑损伤病理过程更加复杂,最终加剧了缺血性脑损伤[32]。
那么从另一个方面,从Gao et al[33]以及Zhang et al[34]发表的多篇文献报道,我们研究人员可得出一些推测性的结论,NSC移植作用所产生的机制的原理,可能是会减少ICH患者中的TCRγδ细胞水平和多种炎症细胞因子的渗透。TCRγδ细胞和IL-17之间本身密不可分的关系,因为TCRγδ细胞可以分泌细胞因子包括IL-17在内的细胞,并且在脑出血发病的病理生理机制的过程中,TCRγδ细胞和IL-17细胞因子这两种生物标志物之间的明确区别在以往的实验研究中几乎没有发现。所以,我们研究人员通过流式细胞仪提取细胞的方法来研究实验,在脑出血血肿周围的组织中发现的 TCRγδ细胞和IL-17 共表达,根据这个结果我们推测出IL-17这种细胞因子在脑出血的患者中神经功能损伤的病理生理机制中发挥主要影响作用,而不是TCRγδ细胞发挥主要作用。
基于这一发现,我们推测可以通过IL-17中和或者其他方式来抑制IL-17细胞因子超载引起ICH患者脑血肿周围组织的过度促炎作用,可能在治疗上减轻ICH的神经毒性作用。而且之前有研究报道,对于ICH患者来说,IL-17中和治疗联合低温治疗相比单纯的抗IL-17细胞因子抗体治疗或者单纯低温治疗有更好的疗效[35]。并且在我们的研究中,我们发现NSCs移植是治疗ICH小鼠实验模型中的一种有效方法,实验研究中我们发现NSCs移植可以促进脑出血小鼠的神经功能恢复。与此同时,IL-17中和与NSCs移植一起可能会增强神经功能的保护作用,说明IL-17中和能有效促进NSCs移植诱导的神经保护作用。然而IL-17在ICH患者的神经功能恢复中的作用机制以及在神经干细胞定向分化中的作用尚未有明确的解释。因此,我们在体外探索了IL-17在神经干细胞发生中的潜在机制,在这里我们隆重的引入了nestin、MAP2和GFAP三种蛋白来区分不同共培养细胞系统中NSCs的分化。Nestin(巢蛋白)是一种生物标志物,它是神经上皮干细胞的一种。GFAP蛋白作为标记物由星形胶质细胞染色来的,MAP2蛋白作为生物标志物同时作为神经元的一种。抗IL-17单克隆抗体在IL-17中和中的应用激发了nsc 中TCRγδ细胞分化的功能。然而,实验中我们并未发现MAP2蛋白的表达中出现这种分化效应。结果表明,IL-17细胞因子可能对NSCs的分化造成深远影响,IL-17细胞因子可以在NSCs向星形胶质细胞和神经元的分化中起抑制作用。抗IL-17单克隆抗体中和IL-17可减轻对星形胶质细胞而非神经元的阻滞作用。虽然中风可以触发NSCs从非活性状态重新分化和增殖活性状态,但已有研究表明,NSCs的迁移和增殖可以被抑制,因此可能是NSCs的迁移和增殖受到抑制,最终导致炎症灶内NSCs的自发性脱髓鞘和神经修复失败,特别强调的是IL-17细胞因子可能对NSCs起矛盾作用。一方面,李等人发表的研究结果得到的结果是,星形胶质细胞分泌的IL-17A可以维持甚至提高脑室下区(SVZ)神经前体细胞(NPCs)的存活率、包括神经元细胞的分化能力、随后的突触形成和脑卒中后的神经功能的恢复。然而另一方面,也有研究人员在研究过程中发现IL-17细胞因子在作为促炎细胞因子起到至关重要的作用,IL-17细胞因子在由NSCs向星形胶质细胞以及少突胶质细胞的增殖和分化中,起到抑制作用,这也可能是研究人员最能接受的结果。在多发性硬化的研究中,有证据显示,在NSCs培养基中加入IL-17这种细胞因子后,发现并观察最终可得出结论,NSCs的数目会随之减少以及形成神经元的能力也随之降低。我们推测IL-17细胞因子在NSCs神经发生中的作用差异可能与IL-17的来源有关。星形胶质细胞起源IL-17细胞因子可能是起到保护作用,然而TCRγδ细胞或者是 Th17细胞起源的 IL-17细胞因子对NSC可能产生毒性作用。此外,目前市场上的抗IL -17单克隆抗体的商业制剂,例如苏金单抗、单抗抗体在感染、慢性炎症反应和自身免疫性疾病中发挥重要作用,这几种抗体制剂已经广泛被用于治疗属于炎症性去变性的银屑病以及类风湿关节炎等免疫型系统疾病; 并且与肿瘤的发生、发展也有密切关系。与此同时,IL-17的中和作用在MS(多发性硬化症)和系统性红斑狼疮的机制研究中也得到了研究,同样我们在脑卒中的神经功能恢复治疗中发现其仍有商业应用的证据。因此我们在这里可以提出一个定义并去证实,我们认为抗IL -17单克隆抗体在ICH患者的神级功能恢复过程的治疗可能是一个有前途的治疗药物,因此我们将会对抗IL -17单克隆抗体开发和药理病理学研究将进一步研究,并且对于我们研究人员来说又是一次重大的挑战。在本研究中,我们提供了证据,表明血清IL-17水平可能与ICH患者神经功能的恢复显著相关。此外我们在研究脑出血模型中,我们曾设想构造动物模型,由于出血性或缺血性脑卒中的类型不易确定,无法控制出血时间、出血量和出血面积,以及消耗大量时间精力的不足。自发性高血压大鼠购买成本高、来源少,也导致了此类脑出血模型在动物实验研究中应用的局限性,与此相比较,通过植入填充物来模拟脑出血模型,目前这种脑出血模型仅用于开颅或微创钻孔引流对颅内血肿清除的研究, 胶原酶诱导脑出血模型,由于该模型机制为脑弥漫性出血,自发性脑出血与之相比出血突然且没有急性占位效应,因此该模型有一定的局限性,不能完全模拟自发性脑出血的病理过程。在小鼠实验模型中,IL-17中和可同时促进神经功能恢复,并进一步抑制神经功能的损伤。IL-17中和的治疗作用可能有助于促进NCS在体外定向分化为星形胶质细胞。我们认为,由于其具有定植特性,星形胶质细胞对脑损伤的反应可能早于免疫细胞。因此,我们将更加关注中风中星形胶质细胞的即时炎症反应。
5 结论
IL-17作为一种促进炎症反应的细胞因子,这种细胞因子具有非常强烈的神经细胞毒性作用,这种神经细胞毒性作用显著影响着sICH患者的神经功能恢复状况。可能是由于IL-17促炎因子抑制了NSC向功能神经细胞的分化从而影响病人预后的功能恢复。实验研究得出IL-17中和的功能会削弱IL-17促炎因子抑制作用,IL-17中和不仅可以加强NSCs的定向分化水平,并且可以显著的促进神经功能的恢复。
本研究由安徽省自然科学基金资助安徽省(1708085MH211)高校拔尖人才基地(gxbjZD11)。
参考文献
[1]. Krishnamurthi RV, Moran AE, Forouzanfar MH, Bennett DA, Mensah GA, Lawes CM, Barker-Collo S, Connor M, Roth GA, Sacco R et al: The global burden of hemorrhagic stroke: a summary of findings from the GBD 2010 study. Global heart 2014, 9(1):101-106.
[2]. Chen CC, Chen X, Li TC, Lin HL, Chu YT, Lee HC, Cheng YK, Chen DC, Tsai SC, Cho DY et al: PG2 for patients with acute spontaneous intracerebral hemorrhage: a double-blind, randomized, placebo-controlled study. Scientific reports 2017, 7:45628.
[3]. Koivunen RJ, Harno H, Tatlisumak T, Putaala J: Depression, anxiety, and cognitive functioning after intracerebral hemorrhage. Acta neurologica Scandinavica 2015, 132(3):179-184.
[4]. Keep RF, Hua Y, Xi G: Intracerebral haemorrhage: mechanisms of injury and therapeutic targets. The Lancet Neurology 2012, 11(8):720-731.
[5] Mac Laughlin B W,Plurad D S,Sheppard W,et al. The impact of intracranial pressure monitoring on mortality after severe traumatic brain injury[J]. Am J Surg,2015, 210( 6) : 1082-1087.
[6] Yuan Q,Wu X,Sun Y,et al. Impact of intracranial pressure monitoring on mortality in patients with traumatic brain injury: a systematic review and meta-analysis[J]. J Neurosurg,2015,122( 3) : 574-587.
[7] Chang C Y,Lin C Y,Chen L C,et al. The predictor of mortality within six-months in patients with spontaneous cerebellar hemorrhage: a retrospective study[J]. PLoS
One,2015,10( 7) : 123-129.
[8] Wu Y T,Li T Y,Chiang S L,et al. Predictors of first week mortality in patients with acute spontaneous cerebellar hemorrhage[J]. Cerebellum,2013,12( 2) : 165-170.
[9].Mendelow AD, Gregson BA, Fernandes HM, Murray GD, Teasdale GM, Hope DT, Karimi A, Shaw MD, Barer DH, investigators S: Early surgery versus initial conservative treatment in patients with spontaneous supratentorial intracerebral haematomas in the International Surgical Trial in Intracerebral Haemorrhage (STICH): a randomised trial. Lancet 2005, 365(9457):387-397.
[10]. Aronowski J, Zhao X: Molecular pathophysiology of cerebral hemorrhage: secondary brain injury. Stroke 2011, 42(6):1781-1786.
[11]. Felberg RA, Grotta JC, Shirzadi AL, Strong R, Narayana P, Hill-Felberg SJ, Aronowski J: Cell death in experimental intracerebral hemorrhage: the “black hole” model of hemorrhagic damage. Annals of neurology 2002, 51(4):517-524.
[12]. Huang FP, Xi G, Keep RF, Hua Y, Nemoianu A, Hoff JT: Brain edema after experimental intracerebral hemorrhage: role of hemoglobin degradation products. Journal of neurosurgery 2002, 96(2):287-293.
[13].Latour LL, Kang DW, Ezzeddine MA, Chalela JA, Warach S: Early blood-brain barrier disruption in human focal brain ischemia. Annals of neurology 2004, 56(4):468-477.
[14].Liebner S, Dijkhuizen RM, Reiss Y, Plate KH, Agalliu D, Constantin G: Functional morphology of the blood-brain barrier in health and disease. Acta neuropathologica 2018, 135(3):311-336.
[15]. Hara K, Yasuhara T, Maki M, Matsukawa N, Masuda T, Yu SJ, Ali M, Yu G, Xu L, Kim SU et al: Neural progenitor NT2N cell lines from teratocarcinoma for transplantation therapy in stroke. Progress in neurobiology 2008, 85(3):318-334.
[16]. Navarro Quiroz E, Navarro Quiroz R, Ahmad M, Gomez Escorcia L, Villarreal JL, Fernandez Ponce C, Aroca Martinez G: Cell Signaling in Neuronal Stem Cells. Cells 2018, 7(7).
[17]. Peruzzotti-Jametti L, Bernstock JD, Vicario N, Costa ASH, Kwok CK, Leonardi T, Booty LM, Bicci I, Balzarotti B, Volpe G et al: Macrophage-Derived Extracellular Succinate Licenses Neural Stem Cells to Suppress Chronic Neuroinflammation. Cell stem cell 2018, 22(3):355-368 e313.
[18]. Ye L, Gao L, Cheng H: Inflammatory Profiles of the Interleukin Family and Network in Cerebral Hemorrhage. Cellular and molecular neurobiology 2018.
[19]. Zhong Q, Zhou K, Liang QL, Lin S, Wang YC, Xiong XY, Meng ZY, Zhao T, Zhu WY, Yang YR et al: Interleukin-23 Secreted by Activated Macrophages Drives gammadeltaT Cell Production of Interleukin-17 to Aggravate Secondary Injury AfterIntracerebral Hemorrhage. Journal of the American Heart Association 2016, 5(10).
[20].Arunachalam P, Ludewig P, Melich P, Arumugam TV, Gerloff C, Prinz I, Magnus T, Gelderblom M: CCR6 (CC Chemokine Receptor 6) Is Essential for the Migration of Detrimental Natural Interleukin-17-Producing gammadelta T Cells in Stroke. Stroke
2017, 48(7):1957-1965.
[21]. Wojkowska DW, Szpakowski P, Glabinski A: Interleukin 17A Promotes Lymphocytes Adhesion and Induces CCL2 and CXCL1 Release from Brain Endothelial Cells. International journal of molecular sciences 2017, 18(5).
[22].Huang L, Zhang L: Neural stem cell therapies and hypoxic-ischemic brain injury. Progress in neurobiology 2018.
[23]. Encinas JM, Fitzsimons CP: Gene regulation in adult neural stem cells. Current challenges and possible applications. Advanced drug delivery reviews 2017, 120:118-132.
[24].Chen L, Qiu R, Li L, He D, Lv H, Wu X, Gu N: The role of exogenous neural stem cells transplantation in cerebral ischemic stroke. Journal of biomedical nanotechnology 2014, 10(11):3219-3230.
[25].Zhang G, Chen L, Chen W, Li B, Yu Y, LinF, Guo X, Wang H, Wu G, Gu B et al: Neural Stem Cells Alleviate Inflammation via Neutralization of IFN-gamma Negative Effect in Ischemic Stroke Model. Journal of
biomedical nanotechnology 2018, 14(6):1178-1188.
[26] Chang SH,Dong C. A novel heterodimeric cytokine consisting
of IL-17 and IL-17F regulates inflammatory responses[J] . Cell
Res,2007, 17(5):435-440.
[27] Reynolds JM,Pappu BP,Peng J,et al. Toll-like receptor 2 signaling in CD4+ T lymphocytes promotes T helper 17 respon
ses and regulates the pathogenesis of autoimmune disease[J] .
Immunity,2010, 32(5):692-702.
[28]. Kolls JK, Linden A: Interleukin-17 family members and inflammation. Immunity 2004, 21(4):467-476.
[29]. Bustamante A, Lopez-Cancio E, Pich S, Penalba A, Giralt D, Garcia-Berrocoso T, Ferrer-Costa C, Gasull T, Hernandez-Perez M, Millan M et al: Blood Biomarkers for the Early Diagnosis of Stroke: The Stroke-Chip Study. Stroke 2017, 48(9):2419-2425.
[30].Liu B, Hu B, Shao S, Wu W, Fan L, Bai G, Shang P, Wang X: CD163/Hemoglobin Oxygenase-1 Pathway Regulates Inflammation in Hematoma Surrounding Tissues after Intracerebral Hemorrhage. Journal of stroke and cerebrovascular diseases : the official journal of National Stroke Association 2015, 24(12):2800-2809.
[31]. Wang XM, Zhang YG, Li AL, Long ZH, Wang D, Li XX, Xia JH, Luo SY, Shan YH: Expressions of serum inflammatory cytokines and their relationship with cerebral edema in patients with acute basal ganglia hemorrhage. European review for medical and pharmacological sciences 2016, 20(13):2868-2871.
[32].Benakis C, Brea D, Caballero S, Faraco G, Moore J, Murphy M, Sita G, Racchumi G, Ling L, Pamer EG et al: Commensal microbiota affects ischemic stroke outcome by regulating intestinal gammadelta T cells. Nature medicine 2016, 22(5):516-523.
[33].Gao L, Lu Q, Huang LJ, Ruan LH, Yang JJ, Huang WL, ZhuGe WS, Zhang YL, Fu B, Jin KL et al: Transplanted neural stem cells modulate regulatory T, gammadelta T cells and corresponding cytokines after intracerebral hemorrhage in rats. International journal of molecular sciences 2014, 15(3):4431-4441.
[34]. Zhang H, Shao B, Zhuge Q, Wang P, Zheng C, Huang W, Yang C, Wang B, Su DM, Jin K: Cross-talk between human neural stem/progenitor cells and peripheral blood mononuclear cells in an allogeneic co-culture model. PloS one 2015, 10(2):e0117432.
[35]. Li Z, Li K, Zhu L, Kan Q, Yan Y, Kumar P, Xu H, Rostami A, Zhang GX: Inhibitory effect of IL-17 on neural stem cell proliferation and neural cell differentiation. BMC immunology 2013, 14:20.
致 谢
时光荏苒,如同白驹过隙,转眼间三年的研究生学习生涯即将结束,在**大学度过的这三年是我人生中最重要的三年。在这期间我逐渐的从一名青涩稚嫩的医学生成长为一名踏入临床实践的医生,我非常自信而有底气地为这份职业感到骄傲与自豪,这都是源于我社会角色的重大转变及社会价值的极大体现,这让我深深体会到医生是阳光下最振奋人心的、最为高尚的职业。我能够走入医生这一行业离不开给予我无私帮助的爸爸妈妈、好友及师长和我的同门师兄弟们,当然也更离不开在临床工作期间给予我信任的每一位患者!在这毕业别离之际,无论有多少不舍和泪水,我仍要对所有的人说一声谢谢!
我要首先想感谢一如既往默默支持我的父母亲。父亲像一座高山时刻耸立在我的身后给予我人生道路上最为坚强的后盾,每当我遇到困难或挫折,父亲永远站在我的身后给予我支持与鼓励,从未改变;母爱如那春日的暖风是我心灵休憩的港湾,让我永远感到温暖与希望的心里充满阳光,使我在一次又一次的挫折中重新振作起来。感谢我最亲爱的爸爸妈妈对我多年来的哺育之恩,感谢你们对我始终如一的信任与支持,给予了我良好的学习和生活环境,支持我选择了阳光下最光荣的职业——医生。
其次我想感谢我最尊敬的导师——xxx教授。他的育人风格学习态度对我影响极大,研究生期间他对我的生活及学习方面给予了无微不至的关怀与照顾,每当我遇到困惑或疑难时,他总是不遗余力的帮我解决。
在我论文撰写过程中xxx教授给予过诸多指导和很大的帮助,在我遇到问题时,老师也总是耐心的为我解惑、悉心指导,使我的思路变得更加清晰和明确,因此才能顺利完成此篇论文,顺利毕业。
此外,由衷的对xxx大学xxx医院、xxx附属医院说一声感谢,是您们给了我作为一名医学生的成长平台。感谢xxx神经外科的xxx教授、xxx教授、
Xxx教授、xxx教授、xxx主任、xxx主任、xxx主任、xxx主任、xxx等老师们,我真诚的感谢以上在工作和学习中为我传道授业解惑的各位老师。
是您们把我从一个懵懂的医学生领入神经外科这个领域从而领略到神经外科学的独特魅力,让我感受到xxx神经外科的最宝贵的亲情、友情和师生情,xxx神经外科是我心中永远的家!愿大家身体健康,工作顺利!
其次还要特别地感谢xxx、xxx博士、xxx博士、xxx博士、xxx博士、xxx博士、xxx博士,感谢xxx、xxx、xxx等师兄、师弟们,还有好室友xxx,感谢你们的宝贵的帮助,希望咱们的师兄弟情永不枯竭!同时感谢xxx附院所有的带教老师,是你们带给我的启发和支持教会我成为一名合格的临床医生!
最后,感谢xxx,是您给了我这个机会让我得以实现职业理想,感谢安医大这个集体、感谢亲爱的室友,无论天涯与海角,同学情永在!

1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:1158,如若转载,请注明出处:https://www.447766.cn/chachong/134734.html,