第1章前言
1.1气候变化对野生动物的影响
近年来因全球气候变暖而引发的气候极端事件越来越多[1-4]。极端天气引发的森林大火更是严重威胁着生活在林中的野生动植物,给人类及野生动物带来了一系列影响。2019年7月以来,澳洲正在经历数十年以来最严重的高温和干旱,9月在澳大利亚东南部爆发的森林大火,持续不断地燃烧了5个多月才熄灭,引发了全世界的关注;在2017年,加拿大不列颠哥伦比亚省也发生了森林大火。
全球气候变暖直接的表现是温度升高[1],自1880年以来,全球平均气温升高了0.85℃[5]。为了应对全球气候变化问题,许多国家通过签订了《联合国气候变化框架公约》、《京都议定书》等公约,来限制温室气体的排放,但是由于海洋和大气循环系统的惯性,气候将会在未来几百年内持续变化[6]。气候变暖引发了许多生态反应[7],给动植物等生物带来了一系列的影响后果,严重影响物种的生存和繁衍,包括:物种分布范围的变化[8]、成体和幼体营养存储减少[9]、全球范围内物种体型的缩小[10]、森林生长季节的延长[11,12]、动植物群落中物种丰度和组成的变化[13,14]。气候变化一直被认为是一些物种灭绝的主要原因[15]。研究表明,气候变化会造成生物物候的改变,导致物种地理分布的变化,加快物种的灭绝速率,改变生态系统的功能[16-20]。全球变暖还带来其它一系列问题,如荒漠化、海平面上升、冰川消退等。卫星数据显示,自19世纪60年代末以来,地球上的积雪和冰面减少了10%[21]。有研究显示,到2100年,海平面可能上升至少1m[22,23]。有研究报道在南极的一些地区,特别是在南极半岛地区和亚南极岛屿上,环境迅速变暖,降水模式也发生变化[24]。在过去的100年里,热带海洋的温度增加了1-2℃[25]。大量环状珊瑚岛被水淹的频率将加快,可达到每年1次。当海水温度长期超过夏季平均值1.08℃以上时,珊瑚礁就会经历全球大规模白化事件,并持续数周。气候变化导致海水温度持续升高,海水变暖、变酸,从而导致热带珊瑚礁的广泛退化[26]。
气候变化会导致物种地理分布的变化,当温度和降水格局发生变化时物种的地理分布范围会随之发生变化[16,18,27]。物种的地理分布取决于其环境耐受性、扩散应变性以及与其他物种的生物相互作用[28]。Kullman的研究发现,温度上升导致瑞典Scandes山脉主要的树种如欧洲云杉(Picea abies)、欧洲赤松(Pinus sylvestris)等海拔极限在20世纪中期上升了至少100m[29]。Lenoir等比较了欧洲西部171种森林植物在20世纪初、20世纪中后期和21世纪初的海拔分布数据,发现气候变暖导致物种的最适分布海拔平均每10年上升29m[30]。研究表明,陆地类群如蝴蝶、昆虫、鸟类、哺乳动物等每十年向较高纬度移动约17km、向较高海拔移动约11m[8],海洋类群如水藻、鱼类等每十年向极地方向移动72km[31-33]。在陆地上,物种在向更凉爽、更高的环境移动[34];在海洋中,它们向水温更低的深水和极地方向移动[6,35]。在英国生活的12种鸟类由于冬季气温升高,分布区向北平均迁移了18.9km[36];加拿大的赤狐(Vulpes vulpes)、北极狐(Vulpes lagopus)由于气温升高,分布区也向北移动[37]。
气候变化会造成生物物候的改变,通常表现为物候期提前[16,19]。气候变化对动植物物候的影响已有大量报道。Sanderson等[38]通过对长途迁徙鸟类的研究发现,从1970年开始的30多年时间里,气候变化不仅影响了鸟类的迁徙时间、迁徙路线,还影响了鸟类繁殖的成功率,因此导致欧洲地区有54%的鸟类种群下降,有的甚至灭绝。Crick等预测到2080年,约有75%的鸟类产卵期将提前[39]。气候变化使得部分水禽由候鸟变为留鸟,不仅影响鸟类在大范围内水平运动,也同样影响山地留居鸟类的垂直运动[40]。除了鸟类以外,在欧洲和北美长期的观测表明其他一些物种的物候也发生了改变,如蝴蝶的羽化时间提前、两栖动物的繁殖期提前以及植物的开花时间提前等。总的来说,自1960年以来,物种春季活动逐渐提前,很可能是对气候变化的反应[21,41]。气候变化对物种的代谢活动也有较大影响,动物是对气候变化最敏感的生物,进食和繁殖行为比较容易受气候变化的影响[42]。
气候变化给野生动物带来严重影响,甚至加快物种的灭绝速率[18],气候变化已经与栖息地丧失等成为物种灭绝的主要因素之一[18,43]。有预测认为温度每升高1℃,就会导致10%的物种灭绝[44],也有学者提出物种的灭绝比例可能会更高,物种的灭绝可能具有多米诺骨牌效应[45]。气候变化对物种造成直接影响和间接影响,直接影响主要是致死温度,温度是影响地球物种最关键的因子之一[40]。Thomas等[46]研究发现,过去几十年时间内气候变化已经使许多物种的丰富度和分布区发生改变,并且预测在不同的气候变化情景下,到2050年墨西哥-澳大利亚区域将有15%-37%的物种灭绝。北美鼠兔(Ochotona princeps)在一些地方的消失或严重减少,标志着气候变化已经引起了物种的灭绝或至少局部灭绝[47];而澳大利亚的啮齿目动物珊瑚裸尾鼠(Melomys rubicola)因海平面上升而已经灭绝[48]。全球变暖对野生动物的最大灭绝风险可能是在热带,因为那里的生物多样性最大。气候变化对全球物种造成了严重影响,是全球物种共同面临的问题。
1.2微气候对野生动物的影响
微气候是指发生在小尺度或局域尺度的气候现象,是局部的温度、湿度、热辐射、气流速度等,其特点是受地表摩擦阻力、土壤类型、地表坡度和方位、植被覆盖、地表水分含量和植物的蒸散作用等的影响形成了近地层特殊气候[49,50]。森林微气候是指在森林植被的影响下形成的特殊小气候[51],受地势、地形、树种组成、树木生长状况、植被覆盖度、土壤含水量、天气状况等一系列因素的影响。而大气候则被认为是地理区或地理带气候条件的总和[52],其气候数据一般通过气象站、卫星探测、雷达探测等获得。
微气候对野生动物的影响已有一些研究。Linda等的研究发现岩雷鸟阿尔卑斯山亚种(Lagopus muta helvetica)的栖息地为其提供了各种各样的微气候。在炎热的夏季,岩石下面有一个稳定的小气候,温度更加凉爽稳定;而岩石顶部的位置更容易受到风和太阳辐射的影响,温度更加炎热。岩雷鸟则会选择岩石下相对凉爽、稳定的微气候,从而减少了炎热的温度给其带来的影响[53]。Patten等的研究表明在较凉爽、较潮湿、较少暴露于风中的微气候中,小草原松鸡(Tympanuchus pallidicinctus)的存活率较高,生存能力更高[54]。Stelzner通过研究草原狒狒(Papio cynocephalus)在各时间段的姿势和环境小气候发现,小气候变量,如空气温度、风速和太阳辐射,在对其行为的影响上呈非线性相互作用。当草原狒狒无法通过选择一个更有利的微环境来减少热应力时,会通过改变它们的姿势和身体方向来对微气候变化作出反应,以尽量减少热应激[55]。研究表明山齿鹑(Colinus virginianus)在栖息地选择时,小气候是其栖息地选择的主要因素,研究结果发现与随机位点相比,栖息地位点提供了较好的条件使得在暴露在高温下的时间较少[56]。
还有些研究表明许多野生动物可以利用有利的微气候环境,如弄蝶(Hesperia comma)、波纹林莺(Sylvia undata)等在温度上升的情况下选择更凉爽的地方,扩大其北方范围限制。同样,也有许多其他例子表明,北方物种在较温暖的南方范围边界上倾向于较冷的微气候地点,如环颈鸫(Turdus torquatus)和一些甲虫等[57,58]。
1.3历史时期气候变化对大熊猫的影响
在第四纪时,整个地球的气候变化的总的来说是逐渐变冷,但历经了冰期与间冰期的交替出现[59]。在冰期,地球覆盖的冰雪从高纬度区域向低纬度区域渐进,温暖潮湿的气候也向低纬度退缩,随即喜暖动植物向赤道附近迁徙。然而,在间冰期间,气候慢慢变暖,大陆冰雪又向高纬度地区撤退,喜暖动植物也向高纬度地区迁移[59]。大熊猫(Ailuropoda melanoeluca)在漫长的系统演化历程中也经历过扩张期和瓶颈期,研究表明历史时期大熊猫的分布变迁与古气候的动荡密切相关,而近代大熊猫的种群数量及分布变迁则主要与人类活动相关[60,61]。
大约800万年前,人类尚处在森林古猿时期,在云南禄丰、元谋等地的热带潮湿沼泽接近森林的地方的出现了大熊猫的直系祖先——始熊猫(Ailuractos),分布于中国西部的云南省[62]。到了更新世初期,出现了比现在大熊猫体型约小一半的小种大熊猫(Ailuropoda micrta),这一时期我国正处于间冰期,温度比较高,并且植被茂密,广泛的分布着亚热带山林,适宜大熊猫生存[60,63,64],小种大熊猫正是在这一时期由发源地向外扩迁,分布于我国西南部以及中部一些地区[65]。随后的过渡成员-大熊猫武陵山亚种(Ailuropoda melanoleuca wulingshanensis)从其化石分布点来看仅在湖南、湖北、广西三省有分布。在更新世的中期,由于云贵高原以及秦岭的抬升,阻挡了西北的干冷季风,使我国的东南部出现了潮湿温暖的气候,成为我国第四纪的最温暖潮湿时期[59],大熊猫巴氏亚种(Ailuropoda melanoleuca baconi)进一步扩散迁移,其分布达到了全盛时期,广布于我国南方、华北,最北已经到达北京周口店,向南可至越南、老挝等地,向东已至东南沿海地带[60]。晚更新世晚期,冰川一次又一次的扩张,气候带南移[66],大熊猫分布范围开始逐渐变小,大理冰期后,气温开始下降,大熊猫开始退缩至云南、贵州、广西等地。全新世,大熊猫范围进一步缩小,到了旧石器时代的化石点已不多[67]。Zhao等[63]对34只野生大熊猫进行了全基因组测序,重建了大熊猫从起源到现在的种群动态历史,结果表明,大熊猫漫长的演化历程经历了两个种群扩张期、两个瓶颈期和两次分化,并分化成三个不同的遗传群体-即秦岭(QIN)、岷山(MIN)、邛崃-大相岭-小相岭-凉山(QXL)。该研究表明大熊猫种群的两次扩张和收缩与古气候变化密切相关,大熊猫种群的扩张是在温暖湿润的间冰期,而冰期的寒冷则导致种群的缩小[63,68]。由此可见,气候变化是数百万年来大熊猫数量波动的主要驱动因素。
自进入新石器时代以来,人类活动可能是造成大熊猫种群衰退的主要原因[69]。距今约2000年时,大熊猫还零星地分布于我国的河南、湖北、湖南、贵州、云南5省[70],栖息地的面积估计在10万km2左右[69]。到18世纪开始,人口数量持续猛增,随着人口的剧增,森林的大量砍伐和农田的垦拓,使大熊猫的分布区迅速缩减,大熊猫不得不向西退缩,现今仅残存于四川盆地向青藏高原过渡的高山峡谷地带[60,69,71,72]。
1.4气候变化对大熊猫未来影响的预测
全球气候变化的直接表现就是温度升高,大熊猫在这样的背景下的未来到底怎样?会不会如地质时期一样随气候变化分布也随之变化?
有许多研究都对气候变化下,大熊猫栖息地适宜性的动态变化做出预测,推测生境整体破碎化程度将增加,种群范围被迫向更高海拔、更高纬度迁移,并且适宜生境面积减小,且未来大熊猫新增适宜生境大多在现有大熊猫分布区外[20,73-78]。吴建国等用CART模型预测了大熊猫在不同气候情境下的分布变化,模拟显示在不同气候情景下全国大熊猫分布范围会有不同程度缩小[79]。Songer等预测到2080年大熊猫现有适宜生境可能丧失60%,生境斑块总数增加,破碎化加剧[74]。Jian等通过物种分布模型预测除秦岭外大熊猫栖息地面积将减少,且总体向北迁移[80]。对邛崃山系的预测也表明大熊猫生境在未来会不同程度地减少[74,81]。刘艳萍在对岷山山系的研究中,认为在未来气候变化背景下,岷山大熊猫适宜生境将减少,66.75%的大熊猫向高海拔扩散,33.25%的大熊猫向低海拔扩散,同时向高纬度区域扩散[82]。在对秦岭山系及大、小相岭山系和凉山山系的预测也显示相似的结果,大熊猫栖息地面积将减少[20,74]。
研究表明,气候变化影响大熊猫的另一方面体现在对大熊猫主食竹的影响上[76]。林下竹子是大熊猫栖息地不可缺少的组成部分,大熊猫将大部分的活动时间用于对竹子的摄食,一只成年大熊猫一天最高可摄食38 kg竹子。然而许多竹种因其有性生殖间隔非常长[83],加上有限的种子散布能力[84]更易受气候变化的影响。关于气候变化下竹子分布变化的预测表明,21世纪中国秦岭的几种主要竹种的分布范围将显著减少[85],到21世纪末,如果竹种不能扩散到现有分布以外的新气候适宜区域,秦岭地区几乎所有的大熊猫栖息地都可能会消失[76]。还有研究表明气候变化将减少大熊猫栖息地竹子物种的多样性,同时,单种竹林的比例将显著增加[77]。
众多模型预测结果都显示大熊猫栖息地面积将减少,分布范围将缩小,大熊猫将向高海拔、高纬度地区移动。但这些研究大都基于大气候环境数据,这种气候模型不能准确地预测复杂的山区气候变化趋势[85]。从第二、第三和第四次全国大熊猫调查结果来看(表1-1),大熊猫种群数量和生境面积都呈增加的趋势。而根据Wei等人2018年通过分析第三、四次大熊猫全国调查数据,结果显示,大约10年间大熊猫分布的平均海拔上移了80m,但并不是因为气候变化所致,也不是因为主食竹分布上移,更可能是由于人类的放牧等干扰因素所致[86]。
表1-1过去30年大熊猫种群数量与栖息地变化
|
调查时间 Survey time |
种群数量(只) Population size |
栖息地面积 (万hm²) Habitat area |
与上次调查栖息地增长比率(%) Growth rate compared with the last survey |
分布县(市、区)(个) Number of county ( city, area ) distributed |
|
| 第二次全国调查(1985-1988) | 1114 | 139.2 | – | 28 | |
|
第三次全国调查 (1999-2002) |
1596 | 230.4 | 65.5 | 33 | |
|
第四次全国调查 (2011-2014) |
1864 | 258 | 11.9 | 37 | |
1.5微气候对大熊猫的影响研究
本文探讨的微气候主要是大熊猫生活的林下层微生境的气候特征。大熊猫是典型的林栖动物,经历的气候是林下微气候,林下微气候可能与外界气候有所差异,森林可能给大熊猫提供了全球气候变化下的庇护所-对外界气候起一定的缓冲作用,大熊猫生境微气候可能较外界气候变化更加小,因此气候变化给大熊猫带来的影响可能得到减缓。因此,探讨微气候对大熊猫的影响很有意义。
大熊猫的活动节律与温度密切相关,偏向于在“最适温度”区域活动[87]。在寒冷的冬季,大熊猫往往向低海拔迁移以抵御冬天的寒冷;在炎热的夏天,大熊猫则向高海拔迁移以躲避夏天的高温。研究指出气象因子对大熊猫季节活动具有重要影响,温度和湿度对其影响最为突出[87]。有学者对大熊猫育幼洞穴的研究指出,大熊猫选择使用的育幼洞穴较未使用过的洞穴更加温暖干燥,以抵御不良气候的影响[88]。由此可见,大熊猫的生存繁衍与气候有着紧密的联系。
那么,如今全球的气候变化到底会给对大熊猫带来怎样的影响呢?前人所做的关于气候变化对大熊猫影响的研究和预测没有将生境复杂性纳入对物种易受气候变化影响的估计,所有温度数据都来自大气候,即气象站、卫星探测等获得的数据,忽略了生境复杂性的一个重要功能–气候缓冲[89,90]。物种生活的小气候可能是高度异质的,与周围的宏观气候有很强的差异[91]。研究表明,微气候异质性可强烈地缓冲了气候变化对物种的影响[92],微气候异质性可以大大降低气候变化所导致的物种灭绝风险[92]。对于在结构复杂的生态系统中的物种来说,这是一个重要的考虑因素,如森林–从地面到冠层它包含许多复杂的结构,其中任何一种都可能使环境温度降低几度[93-95],森林植被可能通过其对太阳辐射、风、降水的拦截和底层湿度的保持的影响来影响微气候[96,97],特定的植被结构特征能够减轻区域气候变化的影响[49,98],森林或能缓冲这些对生物多样性的影响[99]。这种微生境的热缓冲效应可能是防止当前和即将到来的气候变暖的唯一保护措施[94],在气候不断变化的条件下,地上的微生境也将成为野生动物的主要避难所[100,101]。
森林采伐不仅改变动物生境的结构特征(如适宜生境斑块面积和数量减小、生境斑块间的连通性减弱)[102],还改变林分类型、林龄结构等生境质量特征,进而影响动物种群特征。研究表明森林采伐后大熊猫分布与多度的明显减少,在卧龙自然保护区来说,采伐后森林植被至少得花37年时间的恢复,才能恢复到采伐之前的生境状态,恢复得越趋近于原始林,大熊猫分布才逐渐增多[103]。在王朗自然保护区,大熊猫也明显回避有森林采伐和牲畜放牧的生境[104]。森林采伐还对大熊猫主食竹产生了一定的影响,在马边自然保护区,砍伐区相对于非砍伐区郁闭度明显下降;大叶筇竹(Qiongzhuea macrophylla Hsueh)的基径和竹子密度显著偏大,采笋和森林砍伐已经影响到马边大熊猫主食竹的生长[105]。
森林砍伐是否会严重破坏原始林结构,使其冠层结构发生变化,进而影响对气候缓冲的作用?本文以大熊猫生活的微生境为研究点,通过收集原始林和次生林中大熊猫栖息地的温度、相对湿度、光照强度等微气候数据,以探讨大熊猫微生境对气候的缓冲作用。
第2章研究目的、意义及内容
微气候对于生物多样性保存至关重要[106],然而,有关于微气候对大熊猫的影响研究较少。大熊猫是世界生物多样性保护的旗舰种、古老物种的“孑遗”。如今气候变化已对全球物种造成了严重影响,而大熊猫因其栖息地狭窄、繁殖率低、种群规模小、食物单一、分布区破碎化严重等特点[73-76,79,81,85,107,108],可能更易受到气候变化的影响,那么气候变化是否会成为大熊猫新的生存危机?
2.1研究目的
通过调查收集原始林、次生林大熊猫微生境结构数据,以及温度、相对湿度、照度等数据,探讨不同森林类型在结构与微气候上的差异,揭示砍伐对森林结构和微气候缓冲作用的影响,进一步阐明大熊猫偏好原始林的机制及全球气候变化下大熊猫的所面临的挑战。
2.2研究意义
大熊猫是典型的林栖动物,而林下环境复杂多变,这样的微生境中气候可能与外界气候有所差异,而这种差异可能会减缓气候变化对大熊猫带来的影响,成为气候变化背景下大熊猫的“避难所”。
关于大熊猫栖息地的森林微气候的研究未有涉及,本次研究将为今后大熊猫栖息地森林微气候研究提供基础数据。了解不同森林类型的微生境结构差异,有助于探讨森林微气候形成的初步机理。同时,了解不同森林类型微气候的差异,有助于在今后对大熊猫的保护工作中做到有的放矢,有针对性地制定实施保护管理政策。探究大熊猫生存栖息的林下微生境是否对气候有缓冲作用,有助于科学的评估气候变化对大熊猫带来的影响,从而揭示森林在气候变化下对大熊猫所起的重要意义,为下一步保护管理工作指明方向,为今后大熊猫栖息地的恢复以及栖息地的精细化管理提供依据,对未来大熊猫种群的长期续存具有重要意义。
2.3研究内容
(1)通过收集原始林、次生林的微生境数据,分析森林砍伐对微生境结构的影响;
(2)通过比较分析原始林、次生林中大熊猫微生境的温度、相对湿度、照度等气候因素的差异,研究森林砍伐对微气候的影响;
(3)通过比较分析原始林与次生林微气候的变化(差值),探究森林砍伐对微气候缓冲作用的影响,进一步阐述大熊猫偏好原始林的微气候机制,揭示森林环境对气候变化的缓冲效应。
第3章研究区域概况
3.1自然概况
3.1.1地理位置
四川栗子坪国家级自然保护区位于四川盆地西南缘的大渡河中上游,贡嘎山东南面石棉县境内(图3-1),地理位置为102°10´33´´~102°29´07´´E,28°51´02´´~29°08´42´´N,总面积47885 hm2。保护区位于大凉山西部与雅安市交界的小相岭山系,是以保护大熊猫及其栖息的森林生态系统为主的森林及野生动物类型自然保护区。区域内垂直高度落差大,最高海拔点4551m,最低海拔1330m,具有3221m的相对高差。保护区内气候是以亚热带季风气候为基带的山地气候,年均温偏高、年均降水量偏少。
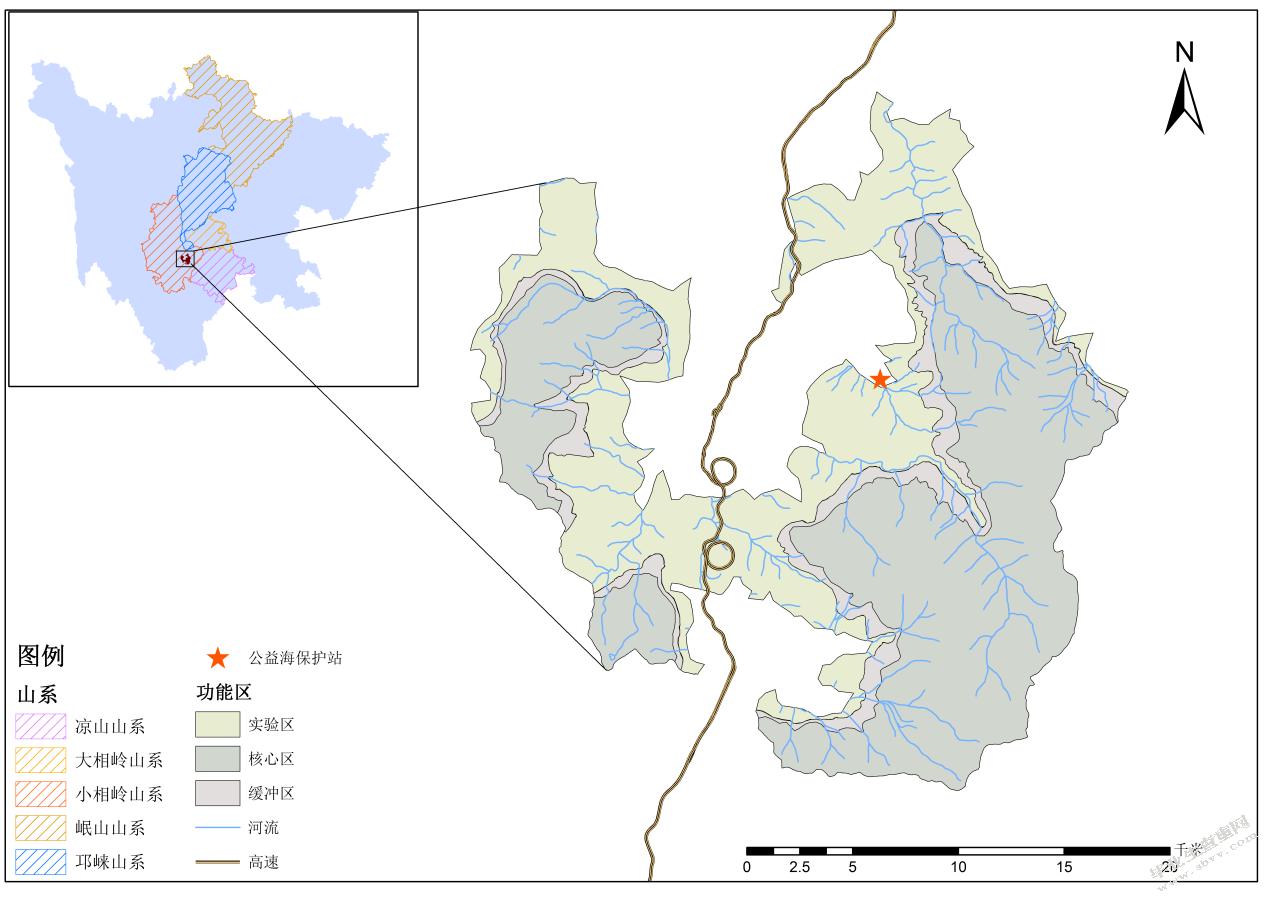

图3-1栗子坪国家级自然保护位置示意图
Fig.3-1 Location of Liziping National Nature Reserve
根据全国第四次大熊猫调查结果(2015),小相岭山系现存大熊猫共30只,是野生大熊猫分布的六大山系中种群数量最少的山系,灭绝概率极高[109]。而栗子坪国家级自然保护区是小相岭大熊猫栖息地的重要组成部分,在保护区内共分布有大熊猫22只,涉及公益海和石灰窑两个局域种群[61](图3-2),栖息地总面积38519.69 km2。
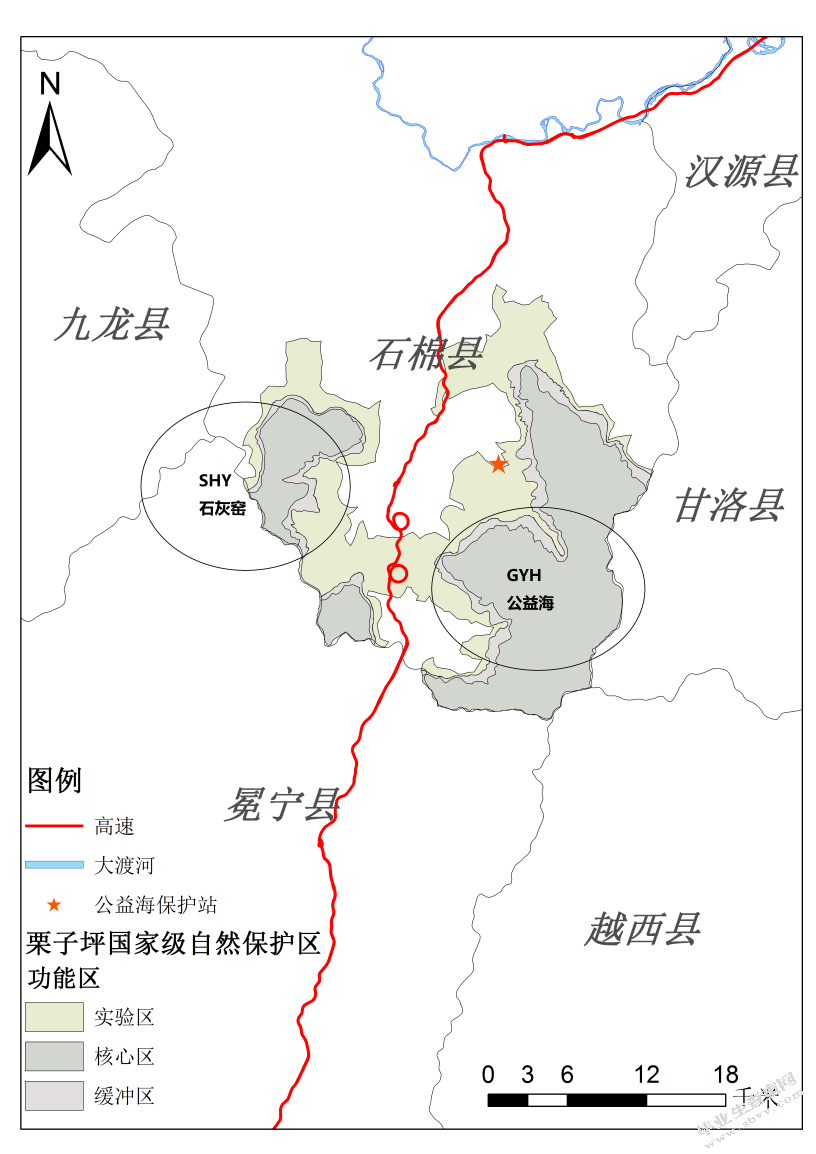
图3-2小相岭山系野生大熊猫局域种群分布
Fig.3-2 Local population distribution of wild giant pandas in the Xiaoxiangling mountains
3.1.2地形、地貌
研究区域位于四川东部平台地与西部地槽的过渡带。由于东部古陆地的长期隆起,该地区属于青藏高原的一个组成部分。地貌以中高山为主,兼有部分低山与河谷阶地。整个地势由西南向东北倾斜,地形切割破碎,起伏跌宕,垂直高差大。总体地貌为峰顶林立,峰谷幽深,具有噶贡山南缘冰蚀山地小区的典型特征。保护区以海拔高度、植被特征等,可进一步划分出高山、中高山、中山和低山四种地貌类型。以地貌特征又可分为:海拔3800m以上为现代季节性冰雪作用及寒冻风化地貌;海拔3500m森林线以上为高山灌木草旬的冰缘地貌;海拔2800m以上为第四纪古冰川作用的残留地貌及坡地灾害地貌;海拔2800m以下为河谷地貌。
3.1.3气候
研究区域气候是以亚热带季风为基带的山地气候。除受地势高低悬殊差异的影响外,大渡河谷对水汽来源和风向、风速影响很大。夏秋受太平洋副热带高压形成的夏季风控制,冬季受喜马拉雅山麓、横断山谷干暖西风控制,春季受青藏高原压脊及西伯利亚高压影响。
区内气候类型属年均温偏高、年均降水量偏少的亚热带季风气候。水热状况垂直变化显著。春夏较长,秋冬略短。气温变幅小,年较差和日较差均不大。年均温约11.7~14.4℃。活动积温较高,植物的生长期较长。
区内降水量800~1250mm,降水分布特点是随海拔升高而递增,冬春干燥,山风强烈。受季风和地形的影响,夏季风控制期主要为沿大渡河进入的东北风,冬季则为来自泸定方向大渡河谷的北风为主。同时,因悬殊的垂直高差及山顶与谷底昼夜温差的变化,山谷风环流现象显著,形成白天由山谷吹向山顶,夜晚由山顶吹向山谷的垂直对流风。
3.1.4水文
研究区域属大渡河一级支流楠垭河流域。区内有公益河、阿鲁伦底河等支流注入楠垭河[110],为典型的树枝状水系的构成之一。区内沟系坡陡谷窄,水流主要靠大气降水沿坡面的径流及高山冰雪融水补给,7~9月为汛期,溪谷多叠水飞瀑。径流内分配不均,沟多坡陡谷窄,小溪河谷多叠水飞瀑,楠垭河全长72 km,流域面积广,水流量充足,沿线建设有多个水利发电站,栗子坪自然保护区在其流域面积内。
3.1.5生物多样性
栗子坪国家级自然保护区内的植坡区划属于亚热带常绿阔叶林区、川东盆地及西南山地常绿阔叶林带、川西南山地偏干性常绿阔叶林带、川西南河山谷原植被地区、大渡河下游高山峡谷植被小区。主要由山地常绿调叶林、亚高山寒温带针叶林、亚高山灌丛、山地针阔叶混交林以及高山灌丛草甸组成[111]。保护区内的建群木种主要由滇青冈(Cyclobalanopsis glaucoides)、多变石栎(Lithocarpus variolosus)等壳斗科类植物组成,山茶科、樟科植物较少。灌木层以茶煎子属(Ribes)、南烛属(Lyonia)等多种落叶成分以及杜鹃属(Rhododendron)为主。
保护区内珍稀动物种类繁多,国家Ⅰ级重点保护的兽类有大熊猫、林麝(Moschus berezovskii)、豹(Panthera pardus)、云豹(Neofelis nebulosa)、羚牛(Budorcas taxicolor)5种,国家Ⅱ级重点保护兽类有小熊猫(Ailurus fulgens)等12种[112]。根据全国第四次大熊猫调查,该保护区内有大熊猫约22只,其中包括2只大熊猫放归个体。从大熊猫的空间分布上看,大熊猫主要分布在保护区内的大林厂-麻麻地-火烧坡范围内的各主要水系附近的针阔混交林及针叶林中。主要以峨热竹(Bashanias panostachya)为食;另外,大熊猫在不同月份还取食海拔2 600 m以下的石棉玉山竹(Yushania lineolata)[113]。
3.2周边社区经济
研究区域周边社区以种植业、养殖业为主。主要粮食有玉米、土豆,经济作物有核桃、花椒等;主要收入靠挖药、核桃和外出务工等:养殖业主要以猪、牛、羊、鸡等品种。保护区周边居民生活条件在不断提高、基础设施建设在不断完善,教育、文化、卫生状况良好,社区居民经济收入有所提高[113]。
但由于长期处于半封闭环境,至今部分边缘山区仍保留着原始的耕作方式,生产力较低,大部分居民观念相当落后。保护区周边社区为彝族居住区,经济落后,交通闭塞,当地居民居住条件简陋,生活艰苦。居民的生活燃料主要以薪柴为主,对森林资源破坏较为严重,导致对野生动植物资源保护难度加大。
3.3历史上森林砍伐概况
自上世纪60年代开始,为提高经济收入,大批森林工厂、伐木厂涌现,大量的森林资源遭到开采和破坏。石棉县的砍伐主要集中在离公路近的区域,可借助汽车运出。另外,河流的沿线也砍伐较多,因为可借助水流的作用将木材运出。后经过1998年“天然林保护工程”、1999年“退耕还林”等保护工程的实施,森林砍伐被全面禁止,森林慢慢恢复。但直到现在,保护区附近居民对森林的依赖性仍然较大,如房屋建筑、柴薪等需求[114]。

图3-3次生林中砍伐后的树桩
Fig.3-3 Tree stumps after felling in secondary forest
第4章研究方法
4.1微生境数据的收集和分析
4.1.1样方的设置
在保护区内大熊猫分布的主要区域,以峨热竹(Bashania spanostachya Yi)分布的起点向上穿越大熊猫活动的生境设置调查样线,分别在原始林和次生林中设置样线10条、18条。原始林是没有经过砍伐、火灾等的原始起源的植被类型;而次生林是砍伐或火灾后自然恢复的植被类型。在样线上设置20m×20m的正方形微生境样方。微生境样方设置地点应满足下列5个条件中的任何一个:
a,峨热竹分布的起止点;
b,遇见大熊猫抓痕、足迹、粪便、采食等痕迹时;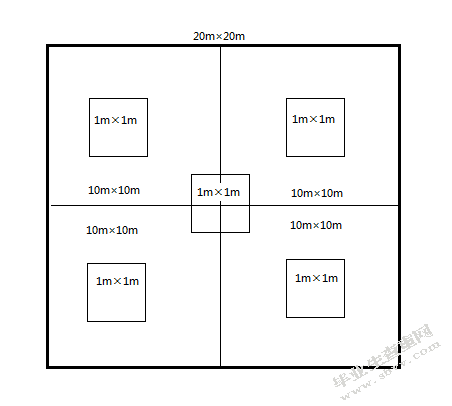
c,植被类型变化区;
d,无上述a—c的特征,但海拔上升或下降了100m或路程约200m时布设样方。
为降低数据在空间上的自相关,以一个样方为准,海拔每上升100m内或行走距离小于200m时不设置样方。在20m×20m正方形大样方中心点及1/4大样方的各10×10m中心设置1个1m2正方形竹子小样方(图4-1)。
在大样方中心点记录生境类型、坡度、坡向、水源距离等变量;以整个大样方为准记录乔木数、乔木郁闭度、乔木胸径、乔木高度;在10m×10m样方内记录灌木株数、灌木高度、灌木盖度;在5个小样方中记录竹子的所有变量,见表4-1。
4.1.2变量设置及定义
野外工作期间,在生境样方内共调查变量20个。各变量定义和测量方法如下(表4-1)。
表4-1各变量的定义和测量方法
| 变量Variables | 定义及测量方法definition and measuring |
|
坡度(0) Gradient |
400m2大样方中心处的坡度,由坡度仪直接读出数字 |
|
水源距离(m) Distance from water |
400m2样方中心与最近终年常流水直线距离 |
|
乔木数 Number of trees |
400m2样方内高度大于5m的乔木数 |
|
乔木郁闭度 Canopy Density(%) |
400m2样方内乔木林冠对地面的覆盖率 |
|
乔木胸径(cm) Diameter at Breast Height(DBH) |
400m2样方内离中心点最近的5颗乔木胸径的平均值 |
|
乔木高度(m) Trees height |
400m2样方内乔木的平均高度 |
|
灌木株数 Shrubs Number |
100m2样方内小于5m的乔木数和灌木丛数 |
|
灌木高度(m) Shrubs Height |
100m2样方内灌木的平均高度 |
|
灌木盖度(%) Shrub Coverage |
100m2样方内灌木对地面的覆盖率(不包括竹子) |
|
竹子盖度(%) Bamboo coverage |
1m2样方内竹子对地面的覆盖率 |
|
死亡竹子数 Bamboo Mortality |
1m2样方内所有死亡竹子株数 |
|
一年生竹株数 Annual number of bamboo |
1m2样方内所有一年生竹的数量 |
|
二年生竹株数 Biennial number of bamboo |
1m2样方内所有二年生竹的数量 |
|
多年生竹株数 Perennial number of bamboo |
1m2样方内所有多年生竹的数量 |
|
一年生竹高(cm) Annual bamboo height |
1m2样方内随机选取5株一年生竹高的平均值 |
|
二年生竹高(cm) Biennial bamboo height |
1m2样方内随机选取5株二年生竹高的平均值 |
|
多年生竹高(cm) Perennial bamboo height |
1m2样方内随机选取5株多年生竹高的平均值 |
|
一年生竹基径(mm) Annual bamboo basal diameter |
1m2样方内随机选取5株一年生竹基径的平均值 |
|
二年生竹基径(mm) Biennial bamboo basal diameter |
1m2样方内随机选取5株二年生竹基径的平均值 |
|
多年生竹基径(mm) Perennial bamboo basal diameter |
1m2样方内随机选取5株多年生竹基径的平均值 |
4.1.3数据分析
对所有变量用Kolmogorov-Smirnov test进行正态分布检验,对满足正态分布的数据采用One-way ANOVA分析,将具有显著差异的变量纳入后面的分析。如果不符合正态分布,则采用平方根转换法进行转换[63],并运用Kruskal-Wallis H Test比较转换后仍然不符合正态分布的数据[49]。
为消除共线性,通过皮尔逊相关性分析(Pearson correlation analysis)分析和斯皮尔曼相关性分析(Spearman correlation analysis)分析正态分布和不具有正态分布的变量,当相关系数小于0.5时,两个变量同时进入后续判别分析;当相关性系数大于0.5时,选择其中更具生物学意义的变量进入后续分析。随后通过逐步判别分析,以找出区分不同生境相对贡献最大的变量。
所有分析均在SPSS20.0中进行。变量数值以平均值士标准差(Mean士SD)表示,显著性水平设为0.05。
4.2微气候数据的收集与分析
4.2.1温湿度计的布设
分别在原始林和次生林各设置3条温湿度计布设的样线,样线之间的水平距离不低于500m。由于该区域大熊猫分布的海拔范围主要在2800-3300m之间[115],因此分别在原始林、次生林中海拔3000m、3100m、3200m处安装温湿度记录仪。
研究中采用的温湿度计型号:HOBO U12-012,生产厂商:Xonset公司。温湿度计安放:距地面1.5m高,统一朝向东南方,固定在乔木或者灌木上,并做防雨保护罩。记录仪时间间隔设置为1h,从2019.04.01号凌晨0:00开始记录。安装如图所示(图4-2)。

图4-2次生林记录仪安装示意图
Fig.4-2 Data logger installation of secondary forest
4.2.2数据分析
将每日微气候数据整理成一组数据,即平均温度、最低温度、最高温度、温度极差、平均相对湿度、最低相对湿度、最高相对湿度、相对湿度极差、平均照度、最小照度、最大照度、照度极差,以便后续处理分析,各变量定义见下表(表4-2)。
表4-2气候变量定义表
|
气候变量 Climate variables |
定义 Definition |
|
平均温度 Mean temperature |
每日2:00、8:00、14:00、20:00四个时刻的平均值 |
|
最低温度 Minimum temperature |
每日温度最低值 |
|
最高温度 Maximum temperature |
每日温度最高值 |
|
温度极差 Range of temperature |
每日温度最高值-每日温度最低值 |
|
平均相对湿度 Mean relative humidity |
每日2:00、8:00、14:00、20:00四个时刻的平均值 |
|
最低相对湿度 Minimum relative humidity |
每日相对湿度最低值 |
|
最高相对湿度 Maximum relative humidity |
每日相对湿度最高值 |
|
相对湿度极差 Range of relative humidity |
每日相对湿度最高值-每日相对湿度最低值 |
|
平均照度 Mean illuminance |
每日2:00、8:00、14:00、20:00四个时刻的平均值 |
|
最小照度 Minimum illuminance |
每日照度最小值 |
|
最大照度 Maximum illuminance |
每日照度最大值 |
|
照度极差 Range of illuminance |
每日照度最大值-每日照度最小值 |
首先通过多因素方差分析(Multivariate analysis of variance)分析森林起源、海拔和时间对气候各变量是否有显著影响,并对有显著影响的单独进行独立样本T检验。然后通过线性混合模型(Linear mixed model)分析森林起源、海拔和时间对气候各变量是否存在交互影响。
所有分析均在SPSS20.0中进行。变量数值以平均值士标准差(Mean士SD)表示,显著性水平设为0.05。
第5章研究结果
于2019年4月至10月进行野外大熊猫微生境调查,共调查微生境样方28个,其中原始林内10个,次生林内18个,填写调查表28份。共布设温湿度计14台,其中原始林中6台,次生林中8台。在野外调查期间,同步收集微生境气候数据,包括时间、温度、相对湿度、光照强度等数据,共3×70413个。
5.1森林砍伐对植被结构的影响
对不同森林起源植被分析结果如表5-1。结果表明:原始林中的坡度、乔木高、乔木胸径、乔木数量、乔木郁闭度、一年生竹基径等变量显著大于次生林。而原始林中的竹子盖度、一年生竹数、二年生竹数、二年生竹高、多年生竹数显著小于次生林。表明原始林和次生林的植被差异较大,原始林内乔木长势优于次生林;而次生林内竹子长势明显优于原始林。
表5-1原始林与次生林各生境变量的单因素方差分析或Kruskal-Wallis H Test检验
|
变量 Variables |
原始林 ( Mean ± SD) |
次生林 ( Mean ± SD) |
t或χ2 | P |
|
坡度 Gradient |
37.50 ± 15.86 | 31.78 ± 9.74 | χ2=7.332 | 0.01* |
|
乔木高度(m) Trees height |
37.30 ± 3.71 | 21.39 ± 6.78 | t=-8.021 | 0.00** |
|
乔木胸径(cm) Diameter at Breast Height(Trees DBH) |
67.20 ± 13.18 | 31.67 ± 9.29 | t=-8.346 | 0.00** |
|
乔木数量 Number of trees |
25.30 ± 3.65 | 11.00 ± 3.42 | t=-10.333 | 0.00** |
|
乔木郁闭度 Trees canopy |
0.67 ± 0.04 | 0.12 ± 0.07 | t=-25.453 | 0.00** |
|
灌木数量 Shrub number |
8.40 ± 0.97 | 7.83 ±2.62 | t=-0.823 | 0.42 |
|
灌木高度(m) Shrub height |
5.90 ± 1.10 | 5.11 ± 1.02 | χ2=3.461 | 0.06 |
|
灌木盖度 Shrub coverage |
0.05 ± 0.02 | 0.06 ± 0.03 | χ2=0.668 | 0.41 |
|
竹子盖度 Bamboo coverage |
0.44 ± 0.07 | 0.77 ± 0.13 | t=8.495 | 0.00** |
|
一年生竹数 Annual number of bamboo |
7.30 ± 1.53 | 12.41 ± 4.62 | t=3.371 | 0.00** |
|
一年生竹高(cm) Annual bamboo height |
225.03 ± 12.20 | 231.37 ± 31.15 | t=0.765 | 0.45 |
|
一年生竹基(mm) Annual bamboo basal diameter |
12.56 ± 0.80 | 11.14 ± 1.44 | t=-2.865 | 0.01** |
|
二年生竹数 Biennial number of bamboo |
7.00 ± 2.47 | 8.98 ± 2.01 | t=2.300 | 0.03* |
|
二年生竹高度(cm) Biennial bamboo height |
218.40 ± 7.74 | 233.90 ± 25.78 | t =2.365 | 0.03* |
|
二年生竹基(mm) Biennial bamboo basal diameter |
12.22 ± 0.92 | 11.49 ± 1.15 | t =-1.726 | 0.10 |
|
多年生竹数 Perennial number of bamboo |
35.40 ± 3.66 | 66.24 ± 16.47 | t =7.613 | 0.00** |
|
多年生竹高度(cm) Perennial bamboo height |
225.02 ± 12.43 | 225.76 ± 34.42 | t =0.082 | 0.94 |
|
多年生竹基(mm) Perennial bamboo basal diameter |
12.00 ± 1.26 | 11.01 ± 1.45 | t =-1.799 | 0.08 |
注:*P<0.05; **P<0.01
当相关系数小于0.5时,则两个变量同时进入判别分析;当变量相关系数大于0.5时,选择其中更具生物学意义的数据进入后续分析。
表5-2 各变量间的相关系数
Tab. 5-2 Correlation coefficient among variables
| 变量 | 乔木胸径 | 乔木数量 | 乔木郁闭度 | 竹子盖度 | 一年生竹数 | 二年生竹数 | 二年生竹高度 | 多年生竹数 |
| 乔木高度 | 0.77 | 0.77 | 0.84 | -0.70 | -0.58 | -0.31 | -0.27 | -0.69 |
| 乔木胸径 | – | 0.73 | 0.87 | -0.74 | -0.57 | -0.55 | -0.24 | -0.80 |
| 乔木数量 | – | – | 0.93 | -0.84 | -0.42 | -0.39 | -0.26 | -0.78 |
| 乔木郁闭度 | – | – | – | -0.86 | -0.59 | -0.48 | -0.22 | -0.85 |
| 竹子盖度 | – | – | – | – | -0.53 | 0.53 | -0.04 | 0.91 |
| 一年生竹数 | – | – | – | – | – | 0.52 | 0.17 | 0.57 |
| 二年生竹数 | – | – | – | – | – | – | -0.22 | 0.65 |
| 二年生竹高度 | – | – | – | – | – | – | – | -0.09 |
最终乔木郁闭度和多年生竹数进入最后的判别函数(如表5-3)。由于逐步判别函数分析只保留相对更大贡献的变量在方程中,因此乔木郁闭度、多年生竹数这2个变量相对于其它变量在判别不同生境的样方上具有更大的贡献力。共建立1个判别函数,对28个微生境样方的总正确判别率达到了100%(表5-4)。
表5-3进入逐步判别函数方程的变量及其判别系数
|
生境变量 Habitat variables |
判别函数系数 Discriminant function |
|
乔木郁闭度 Tree canopy |
1.551 |
|
多年生竹数 Perennial number of bamboo |
0.998 |
表5-4通过逐步判别函数分析对各类生境样方的判别结果
Tab. 5-4 Classification result for habitat plots through Stepwise Discriminant Function Analysis (DFA)
|
森林起源 Forest origin |
原始林 Old-growth forest |
次生林 Secondary forest |
合计 Total |
|
原始林 Old-growth forest |
18 | 0 | 18 |
|
次生林 Secondary forest |
0 | 10 | 10 |
5.2影响气候因子的因素
研究中,就森林类型、海拔以及时间对微生境中各气候因子的影响做多元方差分析,结果如下表(表5-5)。生境起源对平均温度、最低温度、最高温度、温度极差、平均湿度、最小湿度、湿度极差、平均照度、最小照度、最大照度、照度极差有显著影响。海拔对平均温度、最低温度、最高温度、温度极差、最小湿度、湿度极差、平均照度、最大照度、照度极差有显著影响。时间对所有变量都有显著影响。
通过线性混合模型发现,森林类型和海拔对平均温度、最低温度、温度极差、最低湿度、湿度极差、平均照度、最小照度、最大照度、照度极差有交互影响。森林类型和时间对平均温度、最低温度、最大照度、照度极差有交互影响。海拔和时间对平均温度、最低温度有交互影响。森林类型、海拔及时间三者对平均温度、最低温度、最高温度、最小照度、最大照度、照度极差有交互影响。
表5-5森林林型、海拔、时间对各气候变量的影响
|
变量 Variables |
因子 Factors | |||||||||||||
|
森林类型 Forest type |
海拔 Elevation |
时间 Time |
森林类型 * 海拔 Forest type* Elevation |
森林类型 * 时间 Forest type* Time |
海拔 * 时间 Elevation*Time |
森林类型 * 海拔 * 时间 Forest type* Elevation*Time |
||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | F | P | |
|
平均温度 Mean temperature |
4.50 | 0.03 | 133.70 | 0.00 | 1909.17 | 0.00 | 44.02 | 0.00 | 10.62 | 0.16 | 12.086 | 0.05 | 22.09 | 0.00 |
|
最低温度 Minimum temperature |
48.90 | 0.00 | 7.26 | 0.00 | 414.61 | 0.00 | 19.50 | 0.00 | 3.45 | 0.00 | 3.211 | 0.00 | 2.09 | 0.02 |
|
最高温度 Maximum temperature |
15.89 | 0.00 | 19.02 | 0.00 | 87.99 | 0.00 | 2.98 | 0.05 | 1.61 | 0.14 | 0.950 | 0.50 | 1.88 | 0.03 |
|
温度极差 Range of temperature |
70.26 | 0.00 | 10.55 | 0.00 | 155.01 | 0.00 | 19.83 | 0.00 | 1.12 | 0.35 | 0.822 | 0.63 | 1.05 | 0.40 |
|
平均湿度 Mean relative humidity |
4.86 | 0.03 | 0.53 | 0.59 | 209.26 | 0.00 | 0.09 | 0.91 | 0.25 | 0.96 | 0.164 | 1.00 | 0.50 | 0.91 |
|
最低湿度 Minimum relative humidity |
32.88 | 0.00 | 3.52 | 0.03 | 231.08 | 0.00 | 3.61 | 0.03 | 0.83 | 0.55 | 0.20 | 1.00 | 0.44 | 0.95 |
|
最高湿度 Maximum relative humidity |
3.43 | 0.06 | 1.52 | 0.22 | 52.50 | 0.00 | 0.66 | 0.52 | 0.71 | 0.64 | 0.25 | 1.00 | 0.73 | 0.72 |
| 相对湿度极差Range of relative humidity | 47.90 | 0.00 | 5.36 | 0.01 | 213.71 | 0.00 | 5.29 | 0.01 | 1.47 | 0.19 | 0.28 | 0.99 | 0.49 | 0.92 |
|
平均光照强度 Mean illuminance |
137.31 | 0.00 | 135.85 | 0.00 | 40.99 | 0.00 | 136.23 | 0.00 | 1.99 | 0.06 | 0.69 | 0.76 | 1.15 | 0.31 |
|
最小光照强度 Minimum illuminance |
685.13 | 0.00 | 2.07 | 0.13 | 3.13 | 0.01 | 67.15 | 0.00 | 1.47 | 0.18 | 2.83 | 0.00 | 3.01 | 0.00 |
|
最大光照强度 Maximum illuminance |
139.82 | 0.00 | 82.20 | 0.00 | 68.69 | 0.00 | 97.91 | 0.00 | 8.83 | 0.00 | 1.42 | 0.15 | 2.94 | 0.00 |
|
每日光照强度差 Range of daily illuminance |
141.16 | 0.00 | 82.16 | 0.00 | 68.72 | 0.00 | 97.58 | 0.00 | 8.84 | 0.00 | 1.42 | 0.15 | 2.95 | 0.00 |
5.2.1不同森林类型间各气候变量之间的差异
5.2.1.1不同森林类型之间温度的差异
对不同森林类型中温度各指标做独立样本T检验,结果如表5-6。结果显示平均温度、最低温度、最高温度、温度极差在原始林、次生林中均有显著差异。原始林内平均温度(10.17±3.41°C)显著低于次生林平均温度(10.58±3.67°C);最低温度(8.39±3.85°C)也显著高于次生林最低温度(7.45±3.70°C)。而原始林的最高温度(14.10±4.99°C)显著低于次生林最高温度(14.78±5.37°C);原始林温度极差(5.71±4.59°C)显著小于次生林(7.32±5.74°C)。
表5-6不同森林类型之间的温度比较
|
变量 Variables |
因子 Factors | |||||||||||||
|
森林类型 Forest type |
海拔 Elevation |
时间 Time |
森林类型 * 海拔 Forest type* Elevation |
森林类型 * 时间 Forest type* Time |
海拔 * 时间 Elevation*Time |
森林类型 * 海拔 * 时间 Forest type* Elevation*Time |
||||||||
| F | P | F | P | F | P | F | P | F | P | F | P | F | P | |
|
平均温度 Mean temperature |
4.50 | 0.03 | 133.70 | 0.00 | 1909.17 | 0.00 | 44.02 | 0.00 | 10.62 | 0.16 | 12.086 | 0.05 | 22.09 | 0.00 |
|
最低温度 Minimum temperature |
48.90 | 0.00 | 7.26 | 0.00 | 414.61 | 0.00 | 19.50 | 0.00 | 3.45 | 0.00 | 3.211 | 0.00 | 2.09 | 0.02 |
|
最高温度 Maximum temperature |
15.89 | 0.00 | 19.02 | 0.00 | 87.99 | 0.00 | 2.98 | 0.05 | 1.61 | 0.14 | 0.950 | 0.50 | 1.88 | 0.03 |
|
温度极差 Range of temperature |
70.26 | 0.00 | 10.55 | 0.00 | 155.01 | 0.00 | 19.83 | 0.00 | 1.12 | 0.35 | 0.822 | 0.63 | 1.05 | 0.40 |
|
平均湿度 Mean relative humidity |
4.86 | 0.03 | 0.53 | 0.59 | 209.26 | 0.00 | 0.09 | 0.91 | 0.25 | 0.96 | 0.164 | 1.00 | 0.50 | 0.91 |
|
最低湿度 Minimum relative humidity |
32.88 | 0.00 | 3.52 | 0.03 | 231.08 | 0.00 | 3.61 | 0.03 | 0.83 | 0.55 | 0.20 | 1.00 | 0.44 | 0.95 |
|
最高湿度 Maximum relative humidity |
3.43 | 0.06 | 1.52 | 0.22 | 52.50 | 0.00 | 0.66 | 0.52 | 0.71 | 0.64 | 0.25 | 1.00 | 0.73 | 0.72 |
| 相对湿度极差Range of relative humidity | 47.90 | 0.00 | 5.36 | 0.01 | 213.71 | 0.00 | 5.29 | 0.01 | 1.47 | 0.19 | 0.28 | 0.99 | 0.49 | 0.92 |
|
平均光照强度 Mean illuminance |
137.31 | 0.00 | 135.85 | 0.00 | 40.99 | 0.00 | 136.23 | 0.00 | 1.99 | 0.06 | 0.69 | 0.76 | 1.15 | 0.31 |
|
最小光照强度 Minimum illuminance |
685.13 | 0.00 | 2.07 | 0.13 | 3.13 | 0.01 | 67.15 | 0.00 | 1.47 | 0.18 | 2.83 | 0.00 | 3.01 | 0.00 |
|
最大光照强度 Maximum illuminance |
139.82 | 0.00 | 82.20 | 0.00 | 68.69 | 0.00 | 97.91 | 0.00 | 8.83 | 0.00 | 1.42 | 0.15 | 2.94 | 0.00 |
|
每日光照强度差 Range of daily illuminance |
141.16 | 0.00 | 82.16 | 0.00 | 68.72 | 0.00 | 97.58 | 0.00 | 8.84 | 0.00 | 1.42 | 0.15 | 2.95 | 0.00 |
由于时间显著影响了与温度相关的变量,因此逐日将各温度变量绘成曲线图以便直观的看到不同森林类型下各温度变量的差异(图5-1)。原始林和次生林内的平均温度和最低温度总体都呈现出6、7、8三个月份最高,而4、5、9、10、11月份低的趋势;且原始林略高于次生林(图5-1(a),图5-1(b))。而最高温度呈4月、7月、8月最高,5、6、9、10、11月较低的趋势;且原始林中最高温度低于次生林(图5-1(c))。不同森林类型温度极差在不同月份均显现出原始林小于次生林的趋势。
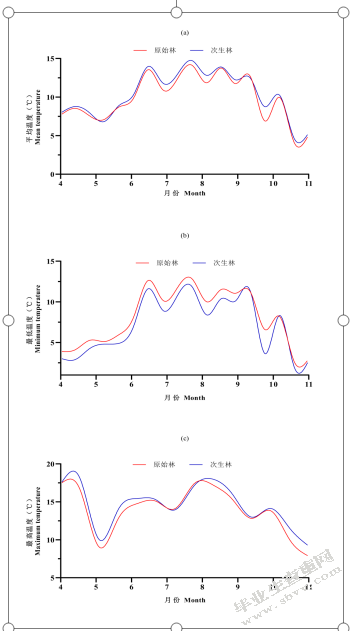
图5-1不同森林类型温度趋势图(a)不同森林类型平均温度;(b)不同森类型最低温度;(c)不同森林类型最高温度
Fig.5-1 Temperature trend of different forest type(a)average temperatures between old-growth forests and secondary forests;(b)minimum temperatures between old-growth forests and secondary forests;(c)maximum temperatures between old-growth forests and secondary forests
进一步比较不同月份内各温度变量在不同森林类型间的差异发现,原始林、次生林内的平均温度在6~9月份有显著差异,均为原始林内平均温度显著低于次生林内平均温度;在4、5、10月份无显著差异,但都略低于次生林(图5-2)。表明原始林在夏季炎热时可以较好的降低林内温度。
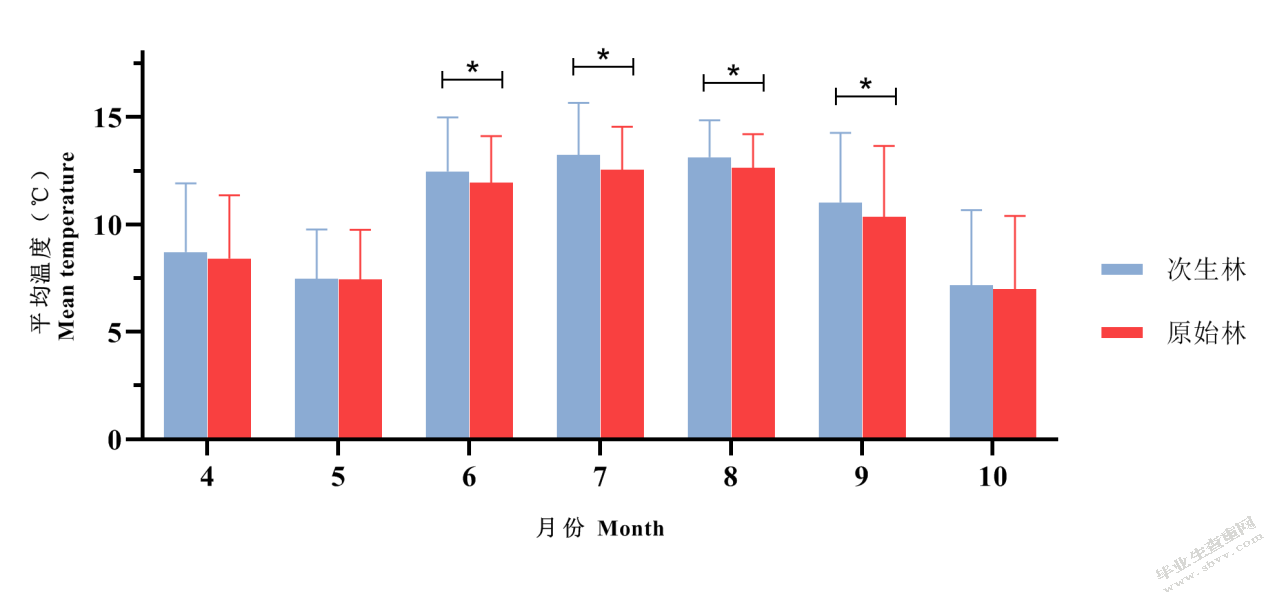
图5-2不同森林类型平均温度比较
Fig.5-2 Comparison of average temperatures between old-growth forests and secondary forests
原始林、次生林内的最低温度在4~9月份有显著差异,原始林内的最低温显著高于次生林。在10月份原始林最低温度略高于次生林,无显著差异(图5-3)。
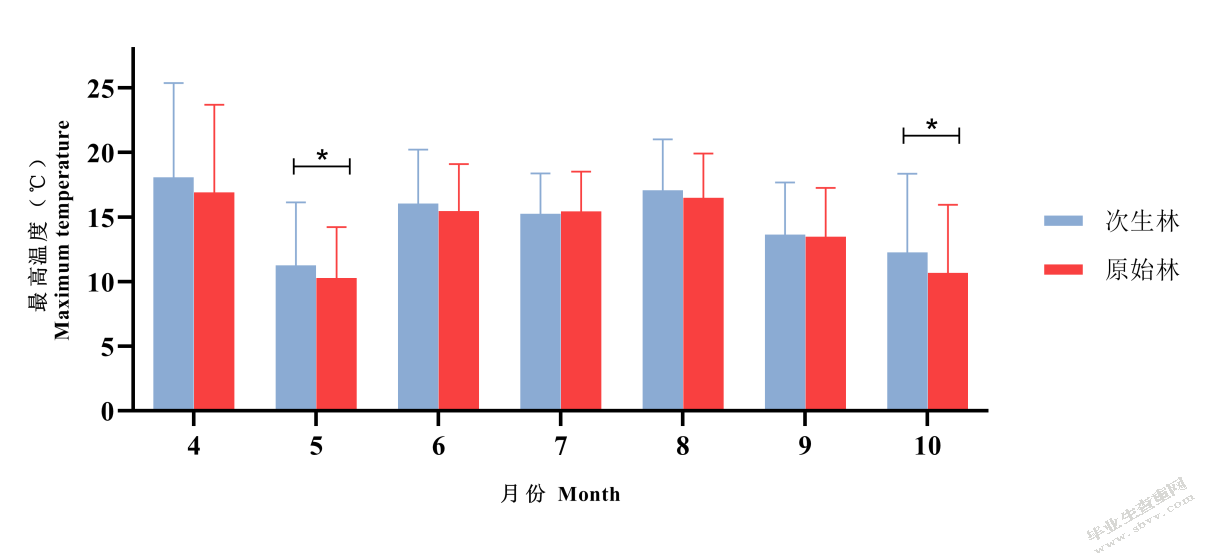
图5-3不同森林类型最低温度比较
Fig.5-3 Comparison of minimum temperatures between old-growth forests and secondary forests
原始林、次生林的最高温度在5、10月份有显著差异,原始林最高温度显著低于次生林。其余月份无显著差异,但原始林内的最高温度均低于次生林内的最高温度(图5-4)。
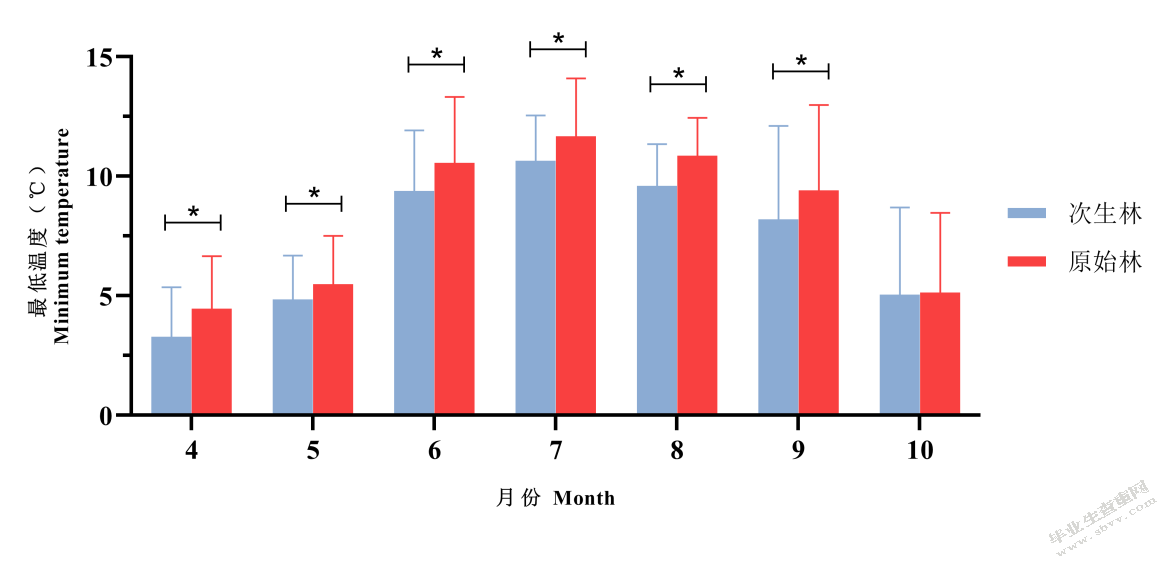
图5-4不同森林类型最低温度比较
Fig.5-4 Comparison of minimum temperatures between old-growth forests and secondary forests
5.2.1.2不同森林类型之间相对湿度的差异
对不同森林类型相对湿度各指标做独立样本T检验,结果如表5-7。结果表明平均湿度、最低湿度、湿度极差在原始林、次生林中均有显著差异,原始林平均相对湿度(95.44±10.79%)高于次生林(94.50±10.67%);最低相对湿度(89.72±19.17%)高于次生林(85.41±22.02%);相对湿度极差(9.03±16.45%)小于次生林(13.59±20.08%)。原始林最高相对湿度(98.75±5.32%)低于次生林(99.00±4.59%),但无显著差异。
表5-7不同森林类型相对湿度湿度比较
|
变量 Variables |
次生林 Secondary forest |
原始林 Old-growth forest |
独立样本T检验 | |
| t | P | |||
|
平均相对湿度 Mean relative humidity(%) |
94.50±10.67 | 95.44±10.79 | -2.26 | 0.02* |
|
最低相对湿度 Minimum relative humidity |
85.41±22.02 | 89.72±19.17 | -5.39 | 0.00** |
|
最高相对湿度 Maximum relative humidity |
99.00±4.59 | 98.75±5.32 | 1.31 | 0.19 |
|
相对湿度极差 Range of relative humidity |
13.59±20.08 | 9.03±16.45 | 6.42 | 0.00** |
将相对湿度各变量绘成折线图以便直观的看到不同森林类型下相对湿度各变量的差异(图5-5)。原始林内平均相对湿度略高于次生林平均相对湿度(图5-5(a))。原始林内最低相对湿度显著高于次生林中(图5-5(b))。最高相对湿度二者并无太大差异(图5-5(c))。
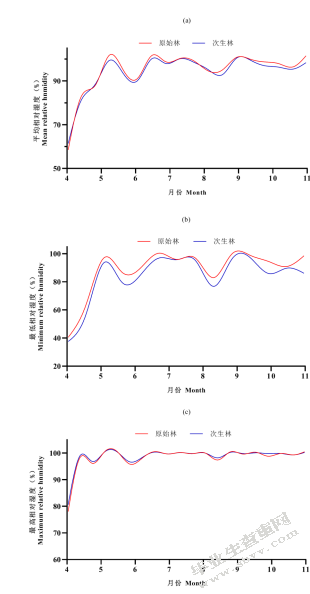
图5-5不同森林类型相对湿度趋势图(a)不同森林类型平均相对湿度;(b)不同森林类型最低相对湿度;(c)不同森林类型最高相对湿度
Fig.5-5 Relative humidity trend of different forest type;(a)average relative humidity between old-growth forests and secondary forests;(b)minimum relative humidity between old-growth forests and secondary forests;(c)maximum relative humidity between old-growth forests and secondary forests
平均相对湿度在6月、7月、8月、9月有显著差异且原始林中平均相对湿度均高于次生林中;其余月份原始林中平均相对湿度略高于次生林(图5-6(a))。原始林中最低相对湿度均高于次生林中,且在4月、5月、6月、8月、9月、10月显著高于次生林中(图5-6(b))。原始林与次生林的最高相对湿度并无显著差异(图5-6(c))。
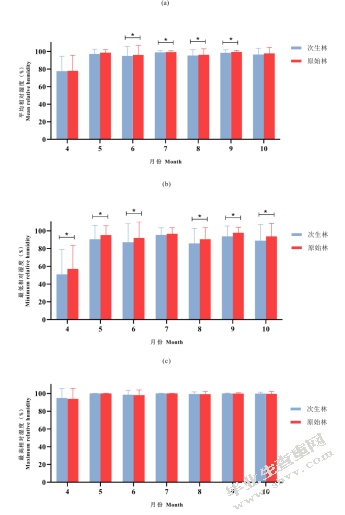
图5-6不同森林类型相对湿度温度比较(a)不同森林类型平均相对湿度;(b)不同森林类型最低相对湿度;(c)不同森林类型最高相对湿度
Fig.5-6 Comparison of relative humidity among different forest type(a)average relative humidity between old-growth forests and secondary forests;(b)minimum relative humidity between old-growth forests and secondary forests;(c)maximum relative humidity between old-growth forests and secondary forests
5.2.1.3不同森林类型照度的差异
太阳福射是地表主要热量来源[116],因此照度也是一项重要指标。对不同森林类型照度各指标做独立样本T检验,结果如表5-8。平均照度、最小照度、最大照度、照度极差在原始林、次生林中均有显著差异。原始林中平均照度(74.35±118.97Lux)显著小于次生林(238.53±453.05Lux),最大照度(456.48±977.93Lux)同样显著小于次生林(1473.56±2767.89Lux)。最小照度(9.39±5.26 Lux)大于次生林(5.17±2.91Lux),有显著差异。
表5-8不同森林类型照度比较
|
变量 Variables |
次生林 Secondary forest |
原始林 Old-growth forest |
独立样本T检验 | |
| t | P | |||
|
平均照度 Mean illuminance(Lux) |
238.53±453.05 | 74.35±118.97 | 13.16 | 0.00** |
|
最低照度 Minimum illuminance(Lux) |
5.17±2.91 | 9.39±5.26 | -24.98 | 0.00** |
|
最高照度 Maximum illuminance(Lux) |
1473.56±2767.89 | 456.48±977.93 | 12.96 | 0.00** |
|
照度极差 Range of illuminance(Lux) |
1468.39±2767.70 | 447.11±976.98 | 13.02 | 0.00** |
将照度各变量绘成折线图以便直观的看到不同森林类型下照度各变量的差异(图5-7)。平均照度及最大照度在原始林和次生林中均呈4月最大,5月开始下降,6月、7月、8月最低,9月开始回升的趋势。且原始林中的平均照度和最大照度均低于次生林中(图5-7(a、c))。最小照度在原始林、次生林中有明显差异且不同月份之间比较稳定,原始林中最小照度略大于次生林,但相比于平均照度和最大照度其差异并不显著(图5-7(b))。
图5-7不同森林类型照度趋势图(a)不同森林类型平均照度;(b)不同森林类型最小照度;(c)不同森林类型最大照度
Fig.5-7 Illuminance trend of different forest type(a)average illuminance between old-growth forests and secondary forests;(b)minimum illuminance between old-growth forests and secondary forests;(c)maximum illuminance between old-growth forests and secondary forests
原始林与次生林中平均照度、最小照度、最大照度在4~10月均有显著差异,且原始林中平均照度、最大照度均显著小于次生林中(图5-8(a、c)),原始林中最小照度略大于次生林中(图5-8(b)),有显著差异。
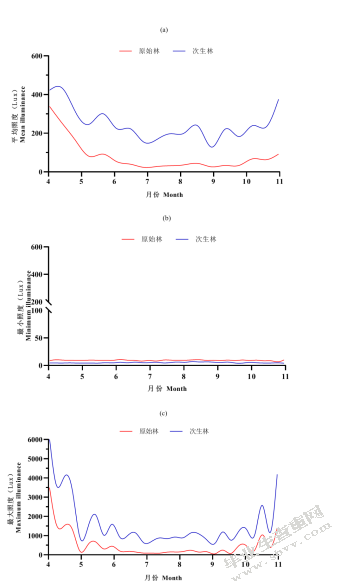
图5-8不同森林类型相照度比较(a)不同森林类型平均照度;(b)不同森林类型最小照度;(c)不同森林类型最大照度
Fig.5-8 Comparison of illuminance among different forest origin(a)average illuminance of different forest origin;(b)minimum illuminance of different forest origin;(c)maximum illuminance of different forest origin
5.2.2不同海拔各气候因子差异
5.2.2.1不同海拔温度的差异
对不同海拔温度做单因素方差分析(表5-9),结果显示海拔对平均温度、最低温度、最高温度、温度极差均有显著影响。平均温度和最低温度呈3000m>3100m>3200m。最高温度和温度极差呈3100 m>3000m>3200m,猜测可能与样本量较少有关;而平均照度和最大照度也同最高温度呈相同趋势(表5-11),为3100m>3000m>3200m,表明了照度与温度内再联系。
表5-9不同海拔温度比较
|
变量 Variables |
海拔(m) | 单因素方差分析 | ||||
| 3000 | 3100 | 3200 | F | P | ||
|
平均温度 Mean temperature (°C) |
10.74±3.45 | 10.44±3.64 | 10.04±3.45 | 7.91 | 0.00** | |
|
最低温度 Minimum temperature (°C) |
8.32±3.75 | 7.78±3.96 | 7.72±3.64 | 5.19 | 0.01* | |
|
最高温度 Maximum temperature (°C) |
14.74±5.19 | 14.93±5.55 | 13.79±4.75 | 13.41 | 0.00** | |
|
温度极差 Range of temperature (°C) |
6.43±5.33 | 7.15±5.81 | 6.07±4.62 | 10.94 | 0.00** | |
海拔越高平均温度越低,3200m分别与3000m、3100m有显著差异(图5-9(a))。最低温度与平均温度有相同趋势,海拔越高温度越低,3000m分别与3100m、3200m有显著差异(图5-9(b))。最高温度则是3100m>3000m>3200m,且3200m分别与3000m、3100m有显著差异(图5-9(c))。温度极差为3100m分别与3000m、3200m有显著差异(图5-9(d))
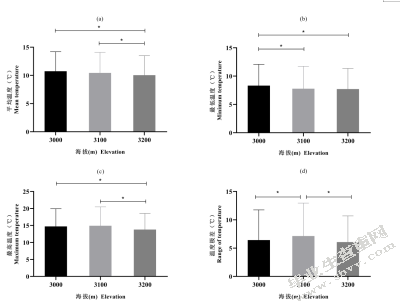 。
。图5-9不同海拔温度的比较(a)不同海拔平均温度;(b)不同海拔最低温度;(c)不同海拔最高温度;(d)不同海拔温度极差
Fig.5-9 Comparison of temperature among different elevation(a)average temperature of different elevation;(b)minimum temperature of different elevation;(c)maximum temperature of different elevation;(d)range of temperature of different elevation
5.2.2.2不同海拔间相对湿度的差异
对不同海拔相对湿度做单因素方差分析(表5-10),结果表明海拔仅对最低相对湿度、相对湿度极差有显著影响,最低相对湿度3000m>3200m>3100m,相对湿度极差3100m>3200m>3000m。对平均相对湿度、最高相对湿度无显著影响。
表5-10不同海拔相对湿度比较
|
变量 Variables |
海拔(m) | 单因素方差分析 | ||||
| 3000 | 3100 | 3200 | F | P | ||
|
平均温度 Mean temperature (°C) |
10.74±3.45 | 10.44±3.64 | 10.04±3.45 | 7.91 | 0.00** | |
|
最低温度 Minimum temperature (°C) |
8.32±3.75 | 7.78±3.96 | 7.72±3.64 | 5.19 | 0.01* | |
|
最高温度 Maximum temperature (°C) |
14.74±5.19 | 14.93±5.55 | 13.79±4.75 | 13.41 | 0.00** | |
|
温度极差 Range of temperature (°C) |
6.43±5.33 | 7.15±5.81 | 6.07±4.62 | 10.94 | 0.00** | |
不同海拔平均温度无显著差异(图5-10(a));最高相对湿度也无显著差异(图5-10(b))。最低相对湿度3100m与3000m、3200m分别有显著差异(图5-10(c))。最低相对湿度在3100m与3000m、3200m分别有显著差异(图5-10(d))。
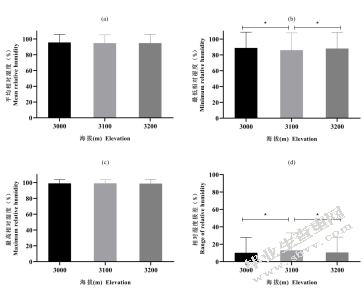
图5-10不同海拔相对湿度的比较(a)不同海拔平均相对湿度;(b)不同海拔最低相对湿度;(c)不同海拔最高相对湿度;(d)不同海拔相对湿度极差
Fig.5-10 Comparison of relative humidity among different elevation(a)average relative humidity of different elevation;(b)minimum relative humidity of different elevation;(c)maximum relative humidity of different elevation;(d)range of relative humidity of different elevation
5.2.2.3不同海拔照度的差异
对不同海拔照度做单因素方差分析(表5-11),海拔对平均照度、最小照度、最大照度、照度极差均有显著影响。平均照度、最大照度及照度极差均为3100m>3000m>3200m,出现与最高温度同样趋势;最小照度为3000m>3100m>3200m。表明海拔对照度有显著影响,不同海拔照度存在差异。
表5-11不同海拔照度比较
|
变量 Variables |
海拔(m) | 单因素方差分析 | ||||
| 3000 | 3100 | 3200 | F | P | ||
|
平均照度 Mean illuminance(Lux) |
89.10±139.91 | 312.34±507.14 | 50.40±86.83 | 180.52 | 0.00** | |
|
最低照度 Minimum illuminance(Lux) |
8.18±6.21 | 7.19±4.33 | 6.4±3.70 | 28.04 | 0.00** | |
|
最高照度 Maximum illuminance(Lux) |
773.05±1526.97 | 1699.77±2997.28 | 412.96±1038.45 | 98.87 | 0.00** | |
|
照度极差 Range of illuminance(Lux) |
764.59±1526.14 | 1692.59±2997.93 | 406.57±1038.45 | 98.78 | 0.00** | |
平均照度呈3100m>3000m>3200m,三个海拔间均有显著差异(图5-11(a))。最低照度随海拔升高而减小,三个海拔间均有显著差异(图5-11(b)),但差异均不超过10Lux。最大照度与平均照度趋势相同,呈3100m>3000m>3200m,三个海拔间均有显著差异(图5-11(c)),最大差异可达1000Lux以上。照度极差3100m>3000m>3200m,三个海拔间均有显著差异(图5-11(d))。
图5-11不同海拔照度的比较(a)不同海拔平均照度;(b)不同海拔最小照度;(c)不同海拔最大照度;(d)不同海拔照度极差
Fig.5-11 Comparison of illuminance among different elevation(a)average illuminance of different elevation;(b)minimum illuminance of different elevation;(c)maximum illuminance of different elevation;(d)range of illuminance of different elevation
5.3森林对气候的缓冲效应
原始林中温度极差(5.71±4.59℃)显著小于次生林(7.32±5.74℃)(图5-12),表明原始林中一天内温度变化范围更小、更稳定,比次生林温度极差小1.61℃,原始林对温度有更显著的缓冲作用。就平均温度而言,原始林在6~9月份显著低于次生林,表明原始林在炎热的夏季能显著地缓冲外界的高温,提供相对凉爽的环境。就最低温度而言,原始林内最低温度(8.39±3.85℃)较次生林最低温度(7.45±3.70℃)高0.94℃,而最高温度则是原始林(14.10±4.99°C)显著低于次生林(14.78±5.37°C)(表5-6)。也就是说,原始林内的温度在最冷的时候较暖,在最热的时候较凉爽。原始林、次生林中温度极差均在4月份最大,原始林日温差达12.45℃,次生林日温差达14.78℃。其余月份原始林、次生林日温差均小于10℃,且原始林温度极差均小于次生林温度极差,表明原始林对温度的缓冲更加明显,其内温度变化较小而更稳定。

图5-12不同森林类型温度缓冲效应
Fig.5-12 Temperature buffer in different forest type
原始林内相对湿度极差(9.03±16.45%)比次生林(13.59±20.08%)低,原始林内的相对湿度变化更小、更加稳定。从相对湿度极差曲线图看原始林内相对湿度极差在同一时间均小于次生林内相对湿度极差(图5-13),表明原始林内相对湿度变化始终较次生林小,也就是说原始林在最潮湿(98.75±5.32%)的时候较次生林(99.00±4.59%)干燥,而在最干燥(89.72±19.17%)的时候较次生林(85.41±22.02%)湿润,原始林对相对湿度有更显著的缓冲作用。原始林、次生林内相对湿度极差在4月最大,原始林相对湿度极达36.65%,次生林中相对湿度极差达43.82%,原始林中相对湿度极差显著低于次生林。其余月份相对湿度极差波动较大,但原始林内相对湿度极差均显著小于次生林。由此可见,原始林对相对湿度有极好的缓冲作用,能保持相对稳定的湿度。
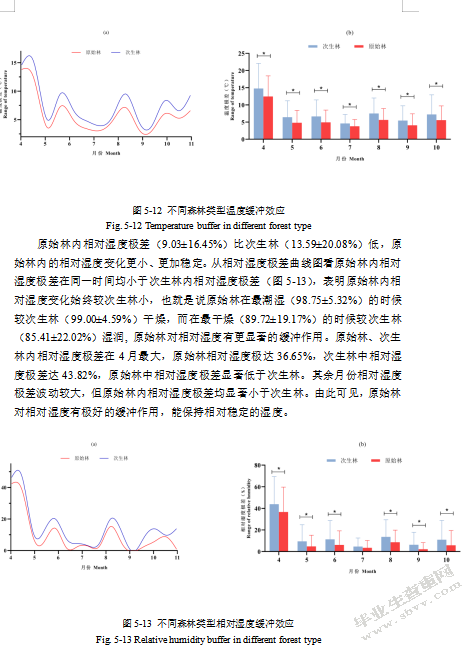
图5-14不同森林类型照度缓冲效应
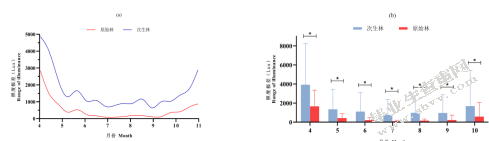
Fig.5-14 Illumination buffer in different forest type
第6章讨论与结论
6.1讨论
6.1.1砍伐对森林结构的影响
大熊猫是一种林栖动物,森林是其生存的基本生境[117]。森林砍伐使得原始林大量丧失、大熊猫栖息地锐减、生境斑块化[113],直接威胁到大熊猫的生存。大熊猫更喜好未经砍伐的原始林,因为其中有较好的育幼、觅食条件,还能更好地躲避天敌、远离干扰[118,119]。
本研究通过对原始林(图6-1(a))、次生林(图6-1(b))植被的分析发现原始林与次生林的差异主要在乔木层和竹子。森林砍伐会改变原有林地的植被结构及自然环境条件,严重改变冠层结构,使得林冠截留能力减弱,对森林生态系统的生态功能过程带来负面的影响[120],从而对大熊猫的分布产生一定的影响。本研究原始林中乔木数量显著大于次生林中乔木数量,高度、胸径、郁闭度也显著大于次生林;洪明生等人在栗子坪自然保护区的研究也表明在原始林中乔木郁闭度、均高、胸径显著大于次生林[113]。原始林一般乔木比较高大,灌木较为稀疏[121]。在王朗大熊猫对不同森林生境的利用的研究也表明,与大熊猫所利用的原始林生境相比较,次生林生境的乔木胸径较小,灌木密度较大[121]。洪明生等人的研究也表明原始林灌木数较少,盖度更小[113]。而本研究中,灌木数量、高及盖度在原始林和次生林中并无显著差异,推测是因为天然保护林等工程的实施,使得次生林中植被得到较好的恢复。
次生林中竹子数量和密度均显著大于原始林。经过恢复,该地竹林恢复较好,竹子长势良好且茂密,绝大部分被竹子覆盖。经砍伐后乔木的郁闭度随之减小,有利于大量的阳光穿透进入林下竹子层,适宜的温度、湿度、光照等有利于竹笋的萌发和竹子生长,因此次生林中竹子密度远远偏大[122]。而原始林中由于冠层的遮挡、光线的不足,竹子数量和长势适中,不及次生林中茂密。研究表明大熊猫更加偏好密度适中的竹林活动和采食[123],本研究的次生林中竹林则过密,不利于大熊猫在其中穿行,原始林竹子密度则适中,也从另一方面证实了大熊猫偏好原始林的原因。
(a)(b)

图6-1当地原始林(a)与次生林(b)
Fig.6-1 Local old-growth forest(a)and secondary forest(b)
6.1.2森林砍伐对微气候的影响
一般而言,森林砍伐会使得林冠层稀疏、光照骤然增强,温湿度变化剧烈,会影响到林内光照强度、空气湿度、温度、土壤养分条件等[120]。本研究中原始林和次生林在温度、湿度、光照等方面均有差异,表明砍伐对森林微气候产生了影响。
森林对气温的影响主要体现在提高极端最低温度、降低极端最高温度、缩小日温度极差、减小温度变化幅度,从而使林内温度保持相对稳定[51]。之前许多研究也表明林地小气候具有降温效应,夏季较凉爽,在冬季气温则不会过低,气温的年较差、日较差较小[124]。本文比较了原始林、次生林的温度指标,发现不同森林类型之间温度差异很大,原始林在保持温度相对稳定上有良好的表现。原始林中平均温度低于次生林中,表明原始林在炎热的夏季能显著地缓冲外界的高温,提供相对凉爽的环境。原始林中最低温度大于次生林,且有极显著差异,在最冷的时候原始林远比次生林温暖。最高温度小于次生林中,有差异显著性。总的来说,原始林中温差较小,在炎热时可以保持相对凉爽而在夜间较冷时则又具保温作用。这可能是因为原始林茂密的冠层、高生物量而有一定的隔热作用[106],在温度较低时阻挡了一部分热量的散失从而林中比外界更加温暖;在温度较高时又阻隔热量进入从而保持凉爽。
与之前有研究表明原始林、次生林林内相对湿度不显著[51],但本研究发现在不同森林起源下相对湿度相差较大。其中原始林平均相对湿度、最低相对湿度大于次生林中,最高相对湿度小于次生林中。推测可能是因为原始林茂密的冠层对湿度有一定的保持作用,而之前的研究在原始林、次生林内无显著差异可能是因为研究地选择的是热带山地雨林,即使是次生林也被茂密的冠层覆盖着。有预测表明大熊猫栖息地气候总体将趋于干暖[108],但就原始林而言,复杂的植被结构对湿度有一定的保持作用,其内湿度仍然保持着一个很高的水平。由于森林砍伐后,原有的森林小气候条件完全改变,冠层减少,对阳光的遮挡作用减弱,阴湿环境发生变化,蓄水能力较强的苔藓和地衣等逐渐衰退[125],次生林中湿度变化范围更加大、更加干燥,相比于原始林其湿度总体偏小。
光照往往是是森林植物生长过程中的主导因子,森林内的照度对植物的生长及森林与大气间的物质和能量的交换起着至关重要的作用[126]。不同森林起源照度差异较大,原始林内平均照度、最大照度小于次生林中,以前有研究也表明原始林内有效辐射小于次生林[51],而最小照度略大于次生林中。可能是因为原始林乔木较多,冠层层次多、叶面积指数大,对太阳辐射有强烈吸收和反射,从而使阳光透过冠层进入林中较少,林内光照强度大大削弱。而次生林由于砍伐冠层大大减少,郁闭度较小,对阳光的吸收和反射作用减弱,竹子也因更多的光照而比原始林长势更好、更茂密。
从本研究的结果来看,温度、相对湿度、照度分别在原始林和次生林中的总体趋势是相似的,表明本次研究数据的有效性。温度随着时间变化有明显的季节性变化(图5-1),在夏季温度偏高,4月、5月、9月、10月偏低,4、5月为春季,9、10月为秋季,温度相对偏低。相对湿度在4月较低,与之前对尖峰岭热带山地雨林的微气候研究结果比较具相似性:空气湿度在5月最低,随后逐渐上升[51]。在进入夏季后雨水丰富,相对湿度相对稳定且偏高。照度随时间变化而变化,4、5月照度较大,可能是因为春季许多树叶正处于生长阶段。6月、7月、8月最低,到了夏季树叶都已经长开,对阳光有阻挡作用。9月逐渐上升,进入秋季树叶慢慢开始掉落,因此林内照度逐渐上升。
6.1.3海拔对微气候的影响
海拔高度是影响气候的重要因子,说明了宏观地形在影响气候方面的重要性[127,128]。本研究的平均温度和最低温度都随着海拔升高而降低,符合规律,但最高温度为3100>3000>3200,推测可能是样本量较少导致的。
不同海拔湿度并无太大差异。海拔不同相对湿度差异不明显,没有显著性。与之前的研究结果相似,不同海拔间空气湿度无明显差异[129]。
照度在不同海拔差异较大,与最高温度呈相同趋势,为3100m最大,3000m次之,3200m最小。最高温度和照度在3100m处最大可能是由于海拔3100m处郁闭度比3000m小,因此温度和照度较海拔3000m处大。虽然在不同海拔照度有所差异,但总体来看差异并不大,可能因为本次研究所选择的海拔梯度较小,且数据记录仪的朝向都相同。
6.1.4原始林–气候变化下大熊猫的庇护所
我们推测原始林对外界气候变化有一定的缓冲作用,原始林茂密的乔木层可以遮挡一部分阳光进入林中进而减少了太阳辐射、阻挡一部分风吹进林中、林中水蒸气散失的更少而保持湿度、降水一部分被截留[106]。因此,原始林中微气候与乔木覆盖较少的次生林有所差异,原始林内微气候更加稳定、更利于大熊猫的生存活动,从另一方面解释了大熊猫偏好原始林的机制。
张合平等通过数据分析表明,森林可以通过林冠层调节环境能量数值和分布来改善系统的小气候条件,降低系统的温度及其变幅[130]。方建民等得出相似的变化规律[131]。Robert的研究表明森林对宏观气候确有缓冲作用,且森林的缓冲能力有显著的季节性变化,夏季的缓冲效应比冬季大[132]。Pieter也曾研究过小气候对林下植物的影响,结果表明小气候效应会减缓气候变暖对森林下层生物的影响,下层植物在小气候的缓冲下能减缓对宏观气候变暖的反应[99]。我们的研究结果也表明,原始林显著地缓冲了外界气候(温度、相对湿度、照度的平均值、最小值、最大值及极差;表5-4、5-5、5-6,图5-1、5-5、5-7)。原始林中平均温度更低、最低温度更温暖,最高温度更凉爽,温度日变化范围比次生林更小。Sarah的研究有着相似的结果,其结果表明与人工林相比,原始林在春季的最高温度较低,并推测可能与原始林的高生物量、高冠层和复杂的垂直结构有关[106]。原始林中平均相对湿度、最低相对湿度更加潮湿,日相对湿度变化范围比次生林中更小。原始林中平均照度、最大照度更小,最小照度比次生林中略大,日照度变化范围比次生林更小。
最低温度对物种的生命产生了深远的影响[133],可能限制着物种分布。本研究结果表明林内的最低温显著高于外界气候最低温,由此降低了最低温给大熊猫带来的限制。森林小气候由冠层封闭调节,其中具有完全封闭冠层的森林表现出较低的日、季节和年变化[134]。我们的研究结果也表明原始林中温度更加稳定,日变化较小。
以前有研究指出由气候变化引起的降雨格局的改变,使部分地区变得更加干旱,加上温度升高蒸散作用加强,使大气及土壤湿度降低[1]。王淑华等研究认为森林具有一定的增湿作用,年平均湿度随高度升高而降低,除冬季外,春夏秋三季湿度变化与降水量均密切相关,并且夏季相对湿度在时间上具有一定的滞后性[135]。本研究结果与王淑华等的研究结果相似,原始林中相对湿度高于次生林中,表明原始林有一定的增湿作用,林内相对湿度并没有受到气候变化的影响。
森林内部更均匀的温度条件是通过树冠层和与水有关的缓冲(即,通过拦截进入的太阳辐射和输出的长波辐射来维持的)[136],森林表现出显著的气候滞后性,并且可以作为对最热敏感的物种的庇护所[106,137,138]。总的来说,与非缓冲环境条件相比,微生境在温度上更凉爽,变化也更小,关键是其减少了大熊猫接触致命温度的机会[139],这表明从树冠到地面的微生境在全球气候变化背景下为大熊猫提供了庇护所。
研究表明大熊猫在选择栖息地时,除竹子这一必然因素外,对原始林的偏好程度大于坡度、地形、海拔等其它因素[119],结合本文的研究,推测大熊猫偏好原始林可能与其气候缓冲作用有一定联系。本研究从微气候角度揭示了原始林对气候的缓冲作用,进而成为气候变化下大熊猫的庇护所。
以往的气候对大熊猫产生影响的研究数据大部分来自气象站、卫星探测等,不能准确地表达森林小气候的特征。而本研究在林内收集其气候数据,更加准确完整。同时在大熊猫栖息地获得如此准确、精细、高质量的微气候数据为今后对大熊猫栖息地微气候的研究提供了科学的环境数据。
6.2结论
1、森林砍伐严重影响了森林结构,减少了高大乔木的存在,郁闭度也变小;然而由于没有上层乔木的对光照等的缓冲作用,林下竹子层的竹子密度显著增加。
2、森林砍伐显著影响了微气候环境。原始林中的平均温度显著低于次生林平均温度;最低温度也显著高于次生林最低温度;而原始林的最高温度显著低于次生林最高温度;温度极差显著小于次生林中。原始林平均相对湿度高于次生林;最低相对湿度高于次生林;原始林最高相对湿度低于次生林;相对湿度极差小于次生林。原始林中平均照度显著小于次生林,最大照度同样显著小于次生林;最小照度显著大于次生林;原始林照度极差显著小于次生林。
3、原始林对气候有一定的缓冲作用,大熊猫所处的微生境内气候温暖湿润,日变化幅度小,气候更加稳定,温度比外界气候更加稳定,对高温和低温都有一定的缓冲;相对湿度也较稳定,波动较小。原始林内微气候条件更利于大熊猫的生存活动。
4、大熊猫更加偏好原始林可能与原始林内较稳定的微气候有关,在全球气候变化的背景下,原始林可能成为大熊猫重要的庇护所。
第7章保护管理建议
全球气候变化日益严峻,在此背景下,原始林内的微生境对气候起着一定的缓冲作用,且林内微气候相对稳定,是大熊猫躲避气候变化的重要庇护所,保护大熊猫栖息地原始林对大熊猫的长期续存意义重大,因此根据本文研究提出以下几点保护管理建议:
(1)加强保护区内大熊猫栖息地原始林的保护工作,成立专门的巡护小组,加强巡护,禁止乱砍乱伐等人为破坏;定期排查,减少火灾等自然灾害的隐患。现有保护区还不能完全覆盖大熊猫栖息地,由于缺少人员管理,保护区外的原始林可能更易受到破坏,因此,不仅要加强各保护区内原始林的保护工作,更要兼顾保护区以外的原始林,守护好大熊猫的家园。
(2)本次调查过程中,虽然未遇见砍伐的情况,但在次生林中挖药的行为依然可见,挖药菜笋等会导致土壤松弛流失,不利于林木的恢复。因此,需要加快次生林的恢复工作,禁止在处在恢复期的次生林内进行林木作业或者采药挖笋等一系列人为活动,以免减缓次生林的恢复过程。
(3)加强大熊猫栖息地周边社区居民的保护意识教育和宣传,增强他们保护意识和主人翁意识。
(4)为保护区周边社区制定行之有效的经济政策,加强社区共管,创新生产生活方式,为保护区周边居民通电通气,改变柴薪取暖的方式,减少居民对树木的依赖;鼓励居民学习新型农业生产技术,鼓励居民“走出大山”,在城镇寻找就业机会,以减少对森林资源的依赖。
(5)加强各保护区、各山系及小种群间原始林的连通性,加快大熊猫廊道建设,加快大熊猫国家公园的建设,实行统一规划管理。
(6)推进大熊猫栖息地微气候在其他山系的研究工作,以便全面掌握大熊猫栖息地的微气候变化动态及规律,有助于科学评估大熊猫在全球气候变化之下所受的影响,从而制定出更精细的栖息地保护和管理政策。
参考文献
[1]Peng S,Li Q,Ren H.Impact of climate change on wildlife[J].Acta Ecologica Sinica,2002,22(7):1153-9.
[2]Abiodun BJ,Lawal KA,Salami AT,et al.Potential influences of global warming on future climate and extreme events in Nigeria[J].Regional Environmental Change,2013,13(3):477-91.
[3]Coma R,Ribes M,Serrano E,et al.Global warming-enhanced stratification and mass mortality events in the Mediterranean[J].Proceedings of the National Academy of Sciences,2009,106(15):6176-81.
[4]Diffenbaugh NS,Singh D,Mankin JS,et al.Quantifying the influence of global warming on unprecedented extreme climate events[J].Proceedings of the National Academy of Sciences,2017,114(19):4881-6.
[5]Pachauri RK,Allen MR,Barros VR,et al.Climate change 2014:synthesis report.Contribution of Working Groups I,II and III to the fifth assessment report of the Intergovernmental Panel on Climate Change[C]:Ipcc;2014.
[6]Pecl GT,Araújo MB,Bell JD,et al.Biodiversity redistribution under climate change:Impacts on ecosystems and human well-being[J].Science,2017,355(6332):eaai9214.
[7]Parmesan C.Ecological and evolutionary responses to recent climate change[J].Annu Rev Ecol Evol Syst,2006,37:637-69.
[8]Chen I-C,Hill JK,Ohlemüller R,et al.Rapid range shifts of species associated with high levels of climate warming[J].Science,2011,333(6045):1024-6.
[9]Derocher AE,Lunn NJ,Stirling I.Polar bears in a warming climate[J].Integrative and comparative biology,2004,44(2):163-76.
[10]Sheridan JA,Bickford D.Shrinking body size as an ecological response to climate change[J].Nature climate change,2011,1(8):401.
[11]Menzel A,Fabian P.Growing season extended in Europe[J].Nature,1999,397(6721):659.
[12]Pretzsch H,Biber P,Schütze G,et al.Forest stand growth dynamics in Central Europe have accelerated since 1870[J].Nature communications,2014,5:4967.
[13]Walther G-R.Community and ecosystem responses to recent climate change[J].Philosophical Transactions of the Royal Society B:Biological Sciences,2010,365(1549):2019-24.
[14]Bellard C,Bertelsmeier C,Leadley P,et al.Impacts of climate change on the future of biodiversity[J].Ecology letters,2012,15(4):365-77.
[15]Maclean I,editor Adapting nature conservation to climate change:the importance of microclimate[C]2018:Open Science Centre,University of Jyväskylä.
[16]Parmesan C,Yohe G.A globally coherent fingerprint of climate change impacts across natural systems[J].Nature,2003,421(6918):37.
[17]Root TL,Price JT,Hall KR,et al.Fingerprints of global warming on wild animals and plants[J].Nature,2003,421(6918):57.
[18]Thomas CD,Franco AM,Hill JK.Range retractions and extinction in the face of climate warming[J].Trends in Ecology&Evolution,2006,21(8):415-6.
[19]Murphy-Klassen HM,Underwood TJ,Sealy SG,et al.Long-term trends in spring arrival dates of migrant birds at Delta Marsh,Manitoba,in relation to climate change[J].The Auk,2005,122(4):1130-48.
[20]Fan J,Li J,Xia R,et al.Assessing the impact of climate change on the habitat distribution of the giant panda in the Qinling Mountains of China[J].Ecological Modelling,2014,274:12-20.
[21]Walther G-R,Post E,Convey P,et al.Ecological responses to recent climate change[J].Nature,2002,416(6879):389.
[22]Vermeer M,Rahmstorf S.Global sea level linked to global temperature[J].Proceedings of the national academy of sciences,2009,106(51):21527-32.
[23]Grinsted A,Moore JC,Jevrejeva S.Reconstructing sea level from paleo and projected temperatures 200 to 2100 AD[J].Climate dynamics,2010,34(4):461-72.
[24]King J,Harangozo S.Climate change in the western Antarctic Peninsula since 1945:observations and possible causes[J].Annals of Glaciology,1998,27:571-5.
[25]Hoegh-Guldberg O.Climate change,coral bleaching and the future of the world's coral reefs[J].Marine and freshwater research,1999,50(8):839-66.
[26]Hoegh-Guldberg O,Mumby PJ,Hooten AJ,et al.Coral reefs under rapid climate change and ocean acidification[J].science,2007,318(5857):1737-42.
[27]Parmesan C.Climate and species'range[J].Nature,1996,382(6594):765-6.
[28]Peterson AT,Soberón J,Pearson RG,et al.Ecological niches and geographic distributions(MPB-49)[M]:Princeton University Press;2011.
[29]Kullman L.20th century climate warming and tree-limit rise in the southern Scandes of Sweden[J].Ambio:A journal of the Human Environment,2001,30(2):72-80.
[30]Lenoir J,Gégout J-C,Marquet P,et al.A significant upward shift in plant species optimum elevation during the 20th century[J].science,2008,320(5884):1768-71.
[31]Poloczanska ES,Brown CJ,Sydeman WJ,et al.Global imprint of climate change on marine life[J].Nature Climate Change,2013,3(10):919-25.
[32]Sorte CJ,Williams SL,Carlton JT.Marine range shifts and species introductions:comparative spread rates and community impacts[J].Global Ecology and Biogeography,2010,19(3):303-16.
[33]Hickling R,Roy DB,Hill JK,et al.The distributions of a wide range of taxonomic groups are expanding polewards[J].Global change biology,2006,12(3):450-5.
[34]Chen I-C,Shiu H-J,Benedick S,et al.Elevation increases in moth assemblages over 42 years on a tropical mountain[J].Proceedings of the National Academy of Sciences,2009,106(5):1479-83.
[35]Dulvy NK,Rogers SI,Jennings S,et al.Climate change and deepening of the North Sea fish assemblage:a biotic indicator of warming seas[J].Journal of Applied Ecology,2008,45(4):1029-39.
[36]Thompson Lo,Mosley-Thompson E,Davis M,et al.“Recent warming”:ice core evidence from tropical ice cores with emphasis on central Asia[J].Global and Planetary Change,1993,7(1-3):145-56.
[37]Hersteinsson P,Macdonald DW.Interspecific competition and the geographical distribution of red and arctic foxes Vulpes vulpes and Alopex lagopus[J].Oikos,1992:505-15.
[38]Sanderson FJ,Donald PF,Pain DJ,et al.Long-term population declines in Afro-Palearctic migrant birds[J].Biological conservation,2006,131(1):93-105.
[39]Crick HQ,Sparks TH.Climate change related to egg-laying trends[J].Nature,1999,399(6735):423-.
[40]马瑞俊,蒋志刚.全球气候变化对野生动物的影响[J].生态学报,2005,11:269-74.
[41]Lozán JL.Climate of the 21st.Century:changes and risks2001.
[42]Post E,Peterson RO,Stenseth NC,et al.Ecosystem consequences of wolf behavioural response to climate[J].Nature,1999,401(6756):905-7.
[43]CBD S,editor Global biodiversity outlook 3[M]2010:Secretariat of the Convention on Biological Diversity Montreal,Quebec,Canada.
[44]Diversity SotCoB.Connecting biodiversity and climate change mitigation and adaptation:Report of the second ad hoc technical expert group on biodiversity and climate change[J].CBD technical series,2009.
[45]Sekercioglu CH,Schneider SH,Fay JP,et al.Climate change,elevational range shifts,and bird extinctions[J].Conservation biology,2008,22(1):140-50.
[46]Thomas CD,Cameron A,Green RE,et al.Extinction risk from climate change[J].Nature,2004,427(6970):145-8.
[47]Stewart JA,Perrine JD,Nichols LB,et al.Revisiting the past to foretell the future:summer temperature and habitat area predict pika extirpations in California[J].Journal of Biogeography,2015,42(5):880-90.
[48]Gynther I,Waller N,Leung LK-P.Confirmation of the extinction of the Bramble Cay melomys Melomys rubicola on Bramble Cay,Torres Strait:results and conclusions from a comprehensive survey in August-September 2014:Queensland Government;2016.
[49]Geiger R.The climate near the ground[M]1957.
[50]Franklin TB.Climates in Miniature:A Study of Micro-Climate and Environment[J].1955.
[51]周璋.海南尖峰岭热带山地雨林小气候特征研究[D]:中国林业科学研究院;2009.
[52]С.П.赫洛莫夫.气候、大气候、地方气候及小气候[J].地理科学进展,1958.
[53]Visinoni L,Pernollet CA,Desmet J-F,et al.Microclimate and microhabitat selection by the Alpine Rock Ptarmigan(Lagopus muta helvetica)during summer[J].Journal of ornithology,2015,156(2):407-17.
[54]Patten MA,Wolfe DH,SHERROD SK,et al.Effects of microhabitat and microclimate selection on adult survivorship of the lesser prairie‐chicken[J].The Journal of wildlife management,2005,69(3):1270-8.
[55]Stelzner JK,Hausfater G.Posture,microclimate,and thermoregulation in yellow baboons[J].Primates,1986,27(4):449-63.
[56]Hiller TL,Guthery FS.Microclimate versus predation risk in roost and covert selection by bobwhites[J].The Journal of wildlife management,2005,69(1):140-9.
[57]Suggitt A,Hodgson J,Maclean I,et al.Microclimate,climate change and wildlife conservation[J].British Wildlife,2014,25(3):162-8.
[58]Gillingham P K,Palmer S C F,Huntley B,et al.The relative importance of climate and habitat in determining the distributions of species at different spatial scales:a case study with ground beetles in Great Britain[J].Ecography,2012,35(9):831-8.
[59]曹伯勋.中国第四纪气候研究及对我国未来气候与环境变化的讨探[J].地质通报,1990,(2):3-17.
[60]张明,袁施彬,张泽钧.大熊猫地史分布变迁初步研究[J].西华师范大学学报(自然科学版),2013,(4):3.
[61]Zhu L,Zhang S,Gu X,et al.Significant genetic boundaries and spatial dynamics of giant pandas occupying fragmented habitat across southwest China[J].Molecular Ecology,2011,20(6):1122-32.
[62]张志和,魏辅文.大熊猫迁地保护理论与实践[M]2006.
[63]Zhao S,Zheng P,Dong S,et al.Whole-genome sequencing of giant pandas provides insights into demographic history and local adaptation[J].Nature Genetics,2013,45(1):67.
[64]邓云霞.历史时期川陕地区大熊猫生境变迁研究[D]:云南大学;2018.
[65]黄万波.大熊猫颅骨,下颌骨及牙齿特征在进化上的意义[J].古脊椎动物学报,031(3):191-207.
[66]周本雄.披毛犀和猛杩象的地理分布,古生态与有关的古气候问题[J].1978.
[67]李涛,赖旭龙,王頠,等.大熊猫的分类与演化综述[J].地质科技情报,2004,(03):40-6.
[68]胡毅,张剑.大熊猫与气候[J].环保科技情报,1993,(3):2-5.
[69]何业恒.大熊猫的兴衰[J].中国历史地理论丛,1998,4:109-23.
[70]Lu Z,Johnson WE,Menotti‐Raymond M,et al.Patterns of Genetic Diversity in Remaining Giant Panda Populations[J].Conservation Biology,2001,15.
[71]文焕然,何业恒.近五千年来豫鄂湘川间的大熊猫[D]1981.
[72]朱立峰,胡义波,张泽钧,等.历史时期人类侵蚀对大熊猫种群的影响[J].Chinese Science Bulletin,2013,58(16):1534-8.
[73]Liu G,Guan T,Dai Q,et al.Impacts of temperature on giant panda habitat in the north Minshan Mountains[J].Ecology and evolution,2016,6(4):987-96.
[74]Songer M,Delion M,Biggs A,et al.Modeling impacts of climate change on giant panda habitat[J].International Journal of Ecology,2012,2012.
[75]Gong M,Guan T,Hou M,et al.Hopes and challenges for giant panda conservation under climate change in the Qinling Mountains of China[J].Ecology and evolution,2017,7(2):596-605.
[76]Tuanmu M-N,Viña A,Winkler JA,et al.Climate-change impacts on understorey bamboo species and giant pandas in China’s Qinling Mountains[J].Nature Climate Change,2013,3(3):249.
[77]Li R,Xu M,Wong MHG,et al.Climate change‐induced decline in bamboo habitats and species diversity:Implications for giant panda conservation[J].Diversity and Distributions,2015,21(4):379-91.
[78]Shen G,Pimm SL,Feng C,et al.Climate change challenges the current conservation strategy for the giant panda[J].Biological Conservation,2015,190:43-50.
[79]吴建国,吕佳佳.气候变化对大熊猫分布的潜在影响[J].环境科学与技术,2009,32(12):168-77.
[80]Jian J,Jiang H,Jiang Z,et al.Predicting giant panda habitat with climate data and calculated habitat suitability index(HSI)map[J].Meteorological Applications,2014,21(2):210-7.
[81]晏婷婷,冉江洪,赵晨皓,等.气候变化对邛崃山系大熊猫主食竹和栖息地分布的影响[J].生态学报,2017,37(07):2360-7.
[82]刘艳萍.气候变化对岷山大熊猫及栖息地的影响[D]:北京:北京林业大学;2012.
[83]Janzen DH.Why bamboos wait so long to flower[J].Annual Review of Ecology and systematics,1976,7(1):347-91.
[84]Taylor AH,Reid DG,Zisheng Q,et al.Spatial patterns and environmental associates of bamboo(Bashania fangiana Yi)after mass-flowering in southwestern China[J].Bulletin of the Torrey Botanical Club,1991:247-54.
[85]Oyler JW,Dobrowski SZ,Ballantyne AP,et al.Artificial amplification of warming trends across the mountains of the western United States[J].Geophysical research letters,2015,42(1):153-61.
[86]Wei W,Swaisgood RR,Dai Q,et al.Giant panda distributional and habitat‐use shifts in a changing landscape[J].Conservation Letters,2018,11(6):e12575.
[87]党超琪,赵凯辉,刘新玉.气象因子对秦岭大熊猫季节性垂直迁徙等活动的影响[J].陕西林业科技,2019.
[88]Wei W,Swaisgood RR,Owen MA,et al.The role of den quality in giant panda conservation[J].Biological conservation,2019,231:189-96.
[89]Suggitt AJ,Gillingham PK,Hill JK,et al.Habitat microclimates drive fine‐scale variation in extreme temperatures[J].Oikos,2011,120(1):1-8.
[90]Williams SE,Shoo LP,Isaac JL,et al.Towards an integrated framework for assessing the vulnerability of species to climate change[J].PLoS biology,2008,6(12):e325.
[91]Potter KA,Arthur Woods H,Pincebourde S.Microclimatic challenges in global change biology[J].Global change biology,2013,19(10):2932-9.
[92]Suggitt AJ,Wilson RJ,Isaac NJ,et al.Extinction risk from climate change is reduced by microclimatic buffering[J].Nature Climate Change,2018,8(8):713.
[93]Shoo LP,Storlie C,Vanderwal J,et al.Targeted protection and restoration to conserve tropical biodiversity in a warming world[J].Global Change Biology,2011,17(1):186-93.
[94]Scheffers BR,Brunner RM,Ramirez SD,et al.Thermal buffering of microhabitats is a critical factor mediating warming vulnerability of frogs in the Philippine biodiversity hotspot[J].Biotropica,2013,45(5):628-35.
[95]Scheffers BR,Phillips BL,Laurance WF,et al.Increasing arboreality with altitude:a novel biogeographic dimension[J].Proceedings of the Royal Society B:Biological Sciences,2013,280(1770):20131581.
[96]Rosenberg NJ,Blad BL,Verma SB.Microclimate:the biological environment[M]:John Wiley&Sons;1983.
[97]Geiger R,Aron RH,Todhunter P.The climate near the ground[M]:Rowman&Littlefield;2009.
[98]Elsen PR,Tingley MW.Global mountain topography and the fate of montane species under climate change[J].Nature Climate Change,2015,5(8):772.
[99]De Frenne P,Rodríguez-Sánchez F,Coomes DA,et al.Microclimate moderates plant responses to macroclimate warming[J].Proceedings of the National Academy of Sciences,2013,110(46):18561-5.
[100]Shoo LP,Storlie C,Williams YM,et al.Potential for mountaintop boulder fields to buffer species against extreme heat stress under climate change[J].International Journal of Biometeorology,2010,54(4):475-8.
[101]Hole DG,Willis SG,Pain DJ,et al.Projected impacts of climate change on a continent‐wide protected area network[J].Ecology letters,2009,12(5):420-31.
[102]Chapin TG,Harrison DJ,Katnik DD.Influence of Landscape Pattern on Habitat Use by American Marten in an Industrial Forest[J].Conservation Biology,1998,12(6):1327-37.
[103]Bearer S,Linderman M,Huang J,et al.Effects of fuelwood collection and timber harvesting on giant panda habitat use[J].Biological Conservation,141(2):0-393.
[104]曾宗永,岳碧松,冉江洪.王朗自然保护区大熊猫对生境的利用[J].四川大学学报(自然科学版),2002,(6):1140-4.
[105]王光磊,周材权.森林砍伐对马边大熊猫主食竹大叶筇竹生长的影响[J].西华师范大学学报(自然科学版),(02):25-8.
[106]Frey SJ,Hadley AS,Johnson SL,et al.Spatial models reveal the microclimatic buffering capacity of old-growth forests[J].Science advances,2016,2(4):e1501392.
[107]McCARTY JP.Ecological consequences of recent climate change[J].Conservation biology,2001,15(2):320-31.
[108]王锐婷,范雄,刘庆,等.气候变化对四川大熊猫栖息地的影响[J].高原山地气象研究,2010,30(04):57-60.
[109]冉江洪,曾宗永,王鸿加,等.四川小相岭山系大熊猫种群及栖息地调查[J].兽类学报,2005,(04):35-40.
[110]马永红,池鸣.四川省栗子坪自然保护区种子植物区系研究[J].西北植物学报,2010,30(06):1240-6.
[111]胡翠华,黄尤优,刘守江,等.四川小相岭山系植被景观的斑块特征分析[J].云南植物研究,2009,31(06):529-36.
[112]李艳红,吴攀文,胡杰.四川栗子坪自然保护区的兽类区系与资源[J].四川动物,2007,(04):841-5.
[113]洪明生,王继成,杨旭煜,等.原始林与次生林中大熊猫微生境结构的比较[J].西华师范大学学报(自然科学版),2012,33(04):356-61.
[114]张君,黄燕.四川栗子坪自然保护区社区经济情况调查[J].西华师范大学学报(自然科学版),(2):5-10.
[115]李杨倩.四川栗子坪国家级自然保护区野生大熊猫种群生存现状[D]:西华师范大学;2019.
[116]王平.西南山地两种阔叶林冠层结构特征的研究[D]:四川农业大学;2016.
[117]冉江洪,曾宗永,王昊,等.大熊猫在原始林和次生林中生境利用的比较研究[J].北京林业大学学报,2004,(04):8-14.
[118]Bearer S,Linderman M,Huang J,et al.Effects of fuelwood collection and timber harvesting on giant panda habitat use[J].Biological Conservation,141(2):0-393.
[119]Zhang Z,Swaisgood RR,Zhang S,等.Old-growth forest is what giant pandas really need[J].Biology Letters,2011,7(3):403-6.
[120]徐崇华,李智宏.森林采伐对群落结构的影响[J].绿色科技,000(004):35-,40.
[121]段利娟,康东伟,王小蓉,等.王朗自然保护区大熊猫原始林与次生林的生境利用比较研究[J].生物学杂志,2014,31(03):49-51.
[122]李杨倩,唐博,周宏,等.历史皆伐林迹地中大熊猫的微生境选择——以栗子坪国家级自然保护区为例[J].西华师范大学学报(自然科学版),39(04):14-20.
[123]张泽钧,胡锦矗.大熊猫生境选择研究[J].西华师范大学学报(自然科学版),(1):18-21.
[124]董慧涵,古伯根,等.白云山风景区林地小气候与环境[J].广州师院学报:自然科学版,000(001):24-33.
[125]林英华,顾海军,隆廷伦,等.森林采伐对平武大熊猫栖息地的影响[J].林业科学,2005,(01):109-15.
[126]李锡泉,田育新,何友军,等.女儿寨小流域植被恢复与重建过程中的小气候效应研究[J].湖南林业科技,032(6):16-8.
[127]Dobrowski SZ.A climatic basis for microrefugia:the influence of terrain on climate[J].Global change biology,2011,17(2):1022-35.
[128]Vanwalleghem T,Meentemeyer R.Predicting forest microclimate in heterogeneous landscapes[J].Ecosystems,2009,12(7):1158-72.
[129]付刚,沈振西.藏北高原不同海拔高度高寒草甸蒸散与环境温湿度的关系[J].中国草地学报,2015,(3).
[130]张合平,田大伦,康文星,等.中国亚热带杉木人工林生态系统的小气候及其形成机制[J].Forestry Studies in China,2001,(02):26-31.
[131]方建民,虞木奎,刘志龙,等.不同农林复合系统小气候日变化特征研究[J].中国园艺文摘,2009,25(09):23-7.
[132]Ewers RM,Banks-Leite C.Fragmentation impairs the microclimate buffering effect of tropical forests[J].PLOS one,2013,8(3):e58093.
[133]Inouye DW.The ecological and evolutionary significance of frost in the context of climate change[J].Ecology Letters,2000,3(5):457-63.
[134]Von Arx G,Graf Pannatier E,Thimonier A,et al.Microclimate in forests with varying leaf area index and soil moisture:potential implications for seedling establishment in a changing climate[J].Journal of Ecology,2013,101(5):1201-13.
[135]王淑华,程春香,孙鹏飞.小兴安岭五营林区气象要素垂直变化规律[J].森林工程,2008,(04):23-6.
[136]Murcia C.Edge effects in fragmented forests:implications for conservation[J].Trends in ecology&evolution,1995,10(2):58-62.
[137]Norris C,Hobson P,Ibisch PL.Microclimate and vegetation function as indicators of forest thermodynamic efficiency[J].Journal of Applied Ecology,2012,49(3):562-70.
[138]Greiser C,Meineri E,Luoto M,et al.Monthly microclimate models in a managed boreal forest landscape[J].Agricultural and forest meteorology,2018,250:147-58.
[139]Scheffers BR,Edwards DP,Diesmos A,et al.Microhabitats reduce animal's exposure to climate extremes[J].Global change biology,2014,20(2):495-503.
下载提示:
1、如文档侵犯商业秘密、侵犯著作权、侵犯人身权等,请点击“文章版权申述”(推荐),也可以打举报电话:18735597641(电话支持时间:9:00-18:30)。
2、网站文档一经付费(服务费),不意味着购买了该文档的版权,仅供个人/单位学习、研究之用,不得用于商业用途,未经授权,严禁复制、发行、汇编、翻译或者网络传播等,侵权必究。
3、本站所有内容均由合作方或网友投稿,本站不对文档的完整性、权威性及其观点立场正确性做任何保证或承诺!文档内容仅供研究参考,付费前请自行鉴别。如您付费,意味着您自己接受本站规则且自行承担风险,本站不退款、不进行额外附加服务。
原创文章,作者:写文章小能手,如若转载,请注明出处:https://www.447766.cn/chachong/15176.html,